FACULTÉ DES SCIENCES
Dr Philippe Reymond
responsable des TP pour médecins
Professeur Jean-Pierre ZRYD
responsable des TP pour biologistes et pharmaciens
Guide de laboratoire à l'usage des étudiants
en biologie, en médecine et en pharmacie
Avec la collaboration du Dr Thierry Diogon
20e édition - 2003 -
FIXATION DE L'AZOTE PAR LES PLANTES
Ces travaux pratiques doivent être considérés comme les compléments nécessaires des cours de Biologie végétale du premier cycle (enseignements propédeutiques : médecins, pharmaciens et biologistes).
Avant chaque description des expériences à faire, de brèves indications théoriques résumeront les problèmes envisagés. Ces notes sommaires ont paru pédagogiquement préférables à de longues explications qui n'auraient fait que répéter, incomplètement, la théorie générale donnée aux cours.
Le choix des thèmes d'expériences n'a pas été facile. Seules des manipulations réalisables en un minimum de temps, nécessitant un appareillage limité et portant sur quelques sujets particuliers, ont été retenues.
Au moment où ils seront programmés, quelques sujets n'auront pas encore nécessairement été traités aux cours. Il est donc indispensable que les étudiants concernés puissent acquérir, à côté des brèves indications théoriques figurant en tête des manipulations, un certain nombre d'informations de base.
Des informations sont disponibles sur le site web du LPC à l’adresse
http://www.unil.ch/lpc/docs/cours1r.htm
ou vous trouverez une bibliographie adéquate et mise à jour (sous la rubrique Travaux Pratiques) – Vous pourrez aussi trouver les liens nécessaires à la consultations de l’encyclopédie « Botany on-line » (en anglais et en allemand)
Physiologie végétale, vol. 1 – Nutrition & vol. 2 - Développement - Heller, Esnault et Lance (Masson, Paris 1998)
Botanique générale - W. Nultsch (DeBoeck-Université, Paris-Bruxelles - 1998) Ouvrage général qui peut trouver son utilité dans tous les cours de botanique et biologie végétale
(édition originale en allemand recommandée aux étudiants maîtrisant cette langue - Thieme Verlag)
Métabolisme des végétaux - G. Richter (Presses Polytechniques et Universitaires Romandes, Lausanne 1993 ; description du livre et commande en ligne possible)
Ouvrage d'un bon niveau qui traite de manière complète les principaux sujets abordés au cours du semestre d'hiver et quelques uns des sujets abordés au semestre d'été - (la dernière édition originale en allemand est recommandée aux étudiants maîtrisant cette langue car elle est plus à jour - Thieme Verlag - 1998).
Botanique – Lüttge, Kluge & Bauer (Lavoisier, Paris 1996)
Biologie - Campbell A. (DeBoeck-Université, Paris-Bruxelles - 1997)
Ouvrage élémentaire dont niveau est plutôt celui de la maturité fédérale. Ce livre peut être cependant utile pour se remettre à jour et pour compléter des lacunes.
Les étudiants seront répartis par groupes. Un tableau de distribution est établi en début de semestre. Les étudiants connaissent donc à l'avance le programme qu'ils auront à suivre et qu'ils devront préparer.
Pour chaque manipulation, et au commencement de la séance de TP, une discussion générale, dirigée par les assistants responsables, portera sur le travail à réaliser durant l'après-midi. Elle permettra aux assistants d'apprécier la préparation de chaque étudiant. Il est absolument inutile que celui-ci essaie de faire les expériences proposées s'il n'a pas préalablement étudié ce qu'il aura à faire concrètement. Il est vivement recommandé à l'étudiant de prendre avec lui ses notes de cours; il pourra ainsi les consulter, les mettre au point et poser à leur sujet toutes les questions utiles.
L'étudiant consignera dans un cahier de laboratoire toutes les données expérimentales concernant les essais réalisés.
Le contenu du journal de laboratoire permettra à chaque sous-groupe de présenter un rapport (sur feuilles séparées) concernant l'une des manipulations effectuées. Le contenu et la forme du rapport (qui peut varier suivant les manipulations) seront indiqués par l'assistant responsable
Les étudiants en médecine présenteront 3 rapports.
Les étudiants en biologie et en pharmacie présenteront 6 rapports.
Le rapport sera remis à l'assistant responsable de la manipulation lors de la séance suivante. Il portera les noms et prénoms des étudiants du sous-groupe, ainsi que leur affiliation (étudiants en médecine A1, A2, B1 ou B2, en biologie, en pharmacie).
L'étudiant se procurera de quoi écrire et dessiner ainsi qu'un cahier quadrillé pour le journal de laboratoire.
Il est vivement conseillé de porter des lunettes et une blouse de laboratoire.
À la fin du semestre d'été, les maîtres-assistants et assistants responsables donneront, pour chaque étudiant, une appréciation sur le travail réalisé.
En ce qui concerne les étudiants en biologie et en pharmacie, l'examen oral peut, à côté du contenu des cours, également porter sur les manipulations effectuées lors des travaux pratiques.
Pour tous les étudiants, les informations du guide font partie intégrante de la matière enseignée.
Lausanne, mars 2003 Dr P. REYMOND
Professeur J.P. ZRYD
La biologie humaine est très influencée par ce que nous mangeons. Beaucoup de substances dont nous avons besoin ne sont pas synthétisées en quantité suffisante par le corps humain ou par sa flore bactérienne. Ces composés, comme les acides gras essentiels, les acides aminés essentiels et les vitamines proviennent de notre alimentation. Les plantes fournissent la part la plus grande et la plus importante de ces composés pour la population mondiale.
" Que ton aliment soit ton médicament " est l'un des principes essentiels de la médecine d'Hippocrate. Depuis plus d'un siècle, les progrès de la connaissance médicale ont en effet établi à plusieurs reprises l'importance d'une alimentation afin de prévenir des pathologies. De nombreux éléments ou composés doivent être présents dans les aliments que nous consommons et cette nécessité n'est pas spécifique à l'alimentation humaine. Inversement. d'autres éléments ou composés doivent être absents de ces mêmes aliments. Il est apparu à la fin du siècle dernier que les aliments consommés en grandes quantités (protéines, graisses. sucres,...) n'étaient pas les seuls nutriments essentiels au bon fonctionnement d'un organisme. Les aliments doivent aussi contenir en faibles quantités, des substances indispensables pour la santé. Ces substances ou micro-nutriments comportent des minéraux et des vitamines.
Les vitamines sont des substances qu'un organisme est incapable d'élaborer lui-même et qui sont indispensables à son développement et à son fonctionnement, et dont l'absence détermine des troubles ou des lésions caractéristiques. Les vitamines sont donc des métabolites essentiels pour certains organismes (hommes, animaux, bactéries) qui doivent les trouver dans leur alimentation. La fonction " vitamine " est donc dépendante de l'organisme considéré. Selon la définition précédente, les acides aminés essentiels pourraient avoir le statut de vitamine, comme cela a été le cas pour les acides gras insaturés, désignés parfois sous le nom de vitamines F.
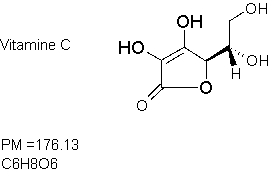
Il est facile de démontrer la présence de vitamine C dans les tissus végétaux par des méthodes biochimiques, dont vous utiliserez l'une d'entre elles dans ces TP. Notez que cette méthode est qualitative et non quantitative – vous pourrez d'ailleurs penser à la façon de rendre cette méthode quantitative. Vous allez utiliser une méthode appelée "empreinte tissulaire". Cette méthode a beaucoup d'applications pour détecter aussi bien de petites que de grandes molécules dans les tissus végétaux.
L'utilisation de l'empreinte tissulaire permet aussi la détection de grandes molécules dans les plantes et en particulier la mise en évidence d'enzymes. Vous allez étudier une peroxydase.
Les peroxydases ont plusieurs fonctions dans les plantes, l'une d'entre elles étant de participer à la synthèse de la lignine, un polymère complexe et très stable de composés phénoliques, localisé dans les parois secondaires (p.ex. dans le xylème).
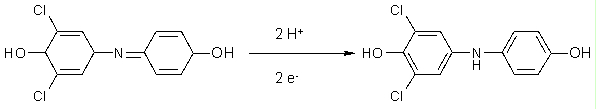
Les peroxydases catalysent la polymérisation de composés phénoliques, dont la structure de base est la suivante:
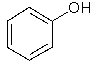
Il y a des milliers de ces composés dans la nature, certains d'entre eux vous étant très familiers, par exemple l'aspirine. Les peroxydases lient beaucoup de composés phénoliques, ce qui fait qu'il est facile d'utiliser des substrats artificiels pour cette enzyme. Le substrat que vous allez employer, la purpurogalline, est un tel exemple de substrat non naturel.
La détection des peroxydases dans les tissus végétaux fournit une excellente démonstration des principes généraux de la mise en évidence d'une enzyme.
Quant des enzymes comme la peroxydase sont couplées à des anticorps spécifiques, nous obtenons un outils puissant et spécifique, la peroxydase est souvent utilisée d'une telle manière. Le chercheur(euse) doit d'abord contrôler le niveau des peroxydases endogènes dans un tissu, entraînant une réaction non spécifique. C'est ce que vous allez faire dans cette manipulation.
L'empreinte tissulaire est une méthode très simple permettant d'obtenir une image physique et chimique d'un tissu sur une surface. Le principe de cette méthode récente est simple : la plupart des composés cellulaires, localisés à la surface de sections fraîchement coupées sur des tissus, peut être transferrée sur une surface adhésive ou absorbante spéciale (p.ex. nitrocellulose) avec peu ou pas de mélange ou de diffusion.
Empreinte physique
|
|
|
|
Vue, au microscope électronique à balayage, de l'image physique laissée sur un film de nitrocellulose par une section de tige de menthe La barre d'échelle représente 40 µm |
Empreinte physique laissée, sur un film sec de colle Sobo, par la surface supérieure d'une feuille de Portulaca. Notez les stomates La barre d'échelle représente 50 µm. |
Empreinte chimique
|
|
|
|
|
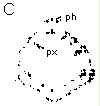
Empreinte d'une section de tige de soja sur de la nitrocellulose et colorée au bleu de toluidine Barre d'échelle : 300 µm ph : phloème primaire px : xylème primaire ca : cambium |
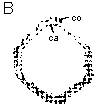
Mise en évidence des protéines riches en hydroxy-proline (HRGPs) par immunocyto-chimie ca : cambium co : cortex |
Mise en évidence des protéines riches en glycine (GRPs) par immunocyto-chimie ph : phloème primaire px : xylème primaire |
Cette technique est actuellement en plein développement. Grâce aux nouveaux supports mis au point (nitrocellulose, nylon, gélatine combinée avec des acides nucléiques,..) et aux méthodes de coloration et d'immunochimie, on peut mettre en évidence et localiser avec précision de nombreuses enzymes et molécules importantes en biologie cellulaire.
Vous aurez l'occasion, dans cette manipulation, de mettre en évidence les structures par coloration du tissu végétal au bleu de toluidine et de localiser une enzyme (la peroxydase) et la vitamine C (acide ascorbique) par empreinte tissulaire.
Bleu de toluidine
Le bleu de toluidine est un colorant non spécifique qui a la particularité de colorer toutes les structures basophiles avec une intensité qui varie selon leur richesse en acides nucléiques et qui est métachromatique avec certaines glycoprotéines riches en eau.
Il colore les polyphénols (lignines, tanins) en vert-turquoise, la cellulose et les pectines en bleu-violet et les protéines en bleu. Il est donc très indiqué si l’on veut rapidement localiser les différents tissus tels xylème, phloème, parenchyme etc. .
Groupes de 2 étudiants : A & B
Prélever l’organe selon les instructions données par les assistants.
A l'aide d'une lame de rasoir, couper 3-4 tranches d'environ 0.5-1 mm.
Placer une petite goutte de colorant (Bleu de toluidine 0.5%) sur une lame.
Poser la coupe sur la goutte de colorant (le colorant ne doit toucher qu'un seul côté de la coupe). Attendre 30 - 60 secondes.
Eliminer l'excès de solvant, sans déplacer la coupe, en mettant en contact le coin d'un morceau de papier-filtre avec le bord de la goutte.
Retourner soigneusement la lame et placez-la sur le support sous la loupe.
Observer l'empreinte en fond noir particulièrement au niveau des vaisseaux de conduction.
On peut détecter de nombreuses enzymes en les faisant agir, directement ou indirectement, sur un substrat chromogène, c'est-à-dire qui se transforme en un composé coloré au cours de la réaction.
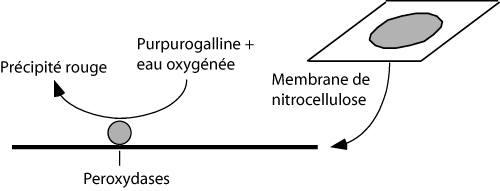
Prélever l’organe selon les instructions données par les assistants.
A l'aide d'une lame de rasoir, couper 3-4 tranches d'environ 2-3 mm d'épaisseur.
Poser soigneusement cette tranche sur le côté lisse d'une feuille de papier de nitrocellulose.
Placer soigneusement une feuille de papier buvard sur la tranche de tissu et presser fermement.
Attention : ne pas déplacer la tranche de tissu lorsque vous pressez.
Enlever délicatement le morceau de buvard sans perturber la tranche de tissu.
En prenant garde de ne pas abîmer le papier de nitrocellulose, enlevez la tranche de tissu.
Placer le morceau de nitrocellulose dans une petite boîte de Pétri contenant 5 ml de solution de purpurogalline 0,06 %.
Faire démarrer la réaction en ajoutant 50 µl d'eau oxygénée H2O2 1% (concentration finale: 0,01 %).
Prélever le morceau de nitrocellulose et le déposer sur une lame de verre.
Examiner où se trouvent localisées les peroxydases.
Localisation de l’acide ascorbique par réduction du nitrate d’argent acide.
Découper une section transversale de 2-3 mm d’épaisseur de la portion centrale d’un pétiole de céleri.
Déposer délicatement la section sur une feuille de papier buvard pour pomper l’excédant de liquide.
Déposer délicatement la même face de la section sur une feuille de nitrocellulose.
Poser un papier buvard par-dessus et presser constamment durant 1min. sans écraser et déformer le tissu.
Plonger la membrane dans la solution de 4% AgNO3 dans 100 mM de tampon Na-acétate (pH 4.0).
Laisser réagir au minimum 10 min.
Observer sous la loupe binoculaire et localiser les zones ayant réagi (contenant de l’acide ascorbique)
Les plantes ont leur propre système de défense qui est différent de celui du monde animal. En règle générale, la défense d'une plante n'est pas assurée par des cellules spécialisées. Toutes les cellules végétales peuvent produire un arsenal de défenses chimiques qui leur permettent de combattre les agents pathogènes.
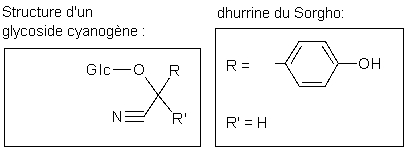
Certaines de ces défenses sont constitutives et sont donc fabriquées continuellement, même en l'absence de pathogènes. Ces composés de faibles poids moléculaires sont appelés phytoanticipines et incluent un grand nombre de molécules bien connues comme des alcaloïdes (nicotine, morphine), des terpènes (l'arôme des épices) ou des composés phénoliques (tannins). Certaines plantes fabriquent une classe de phytoanticipines appelées "glycosides cyanogéniques".
Ces petites molécules sont assez instables et libèrent de l'acide cyanhydrique lorsqu'elles sont dégradées. En effet, lors de l'attaque par un pathogène ou par un herbivore, les cellules sont détruites et les composés cyanogéniques entrent en contact avec des glycosidases dont l'action conduit à la libération de HCN, puissant inhibiteur de la respiration aérobie. Les glycosides cyanogéniques se trouvent dans de nombreuses espèces et dans différents types d'organes (feuilles, fleurs, graines, ...). Les concentrations moyennes varient de 0.5 à 5 µmol/g de poids frais dans les feuilles et de 0.1 à 0.6 µmol/ g dans les graines.
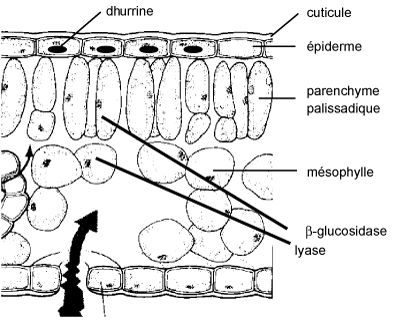
Stockage des glucosides cyanogènes dans la feuille de sorgho:
Composés cyanogéniques dans quelques espèces
|
Espèce |
Organe |
Composé cyanogénique |
|
Lotus corniculatus |
feuille |
linamarine |
|
Manihot esculenta |
tubercule |
linamarine |
|
Prunus laurocerasus |
feuille |
prunasine |
|
Prunus lusitanica |
fruit |
amygdaline |
|
Ranunculus repens |
feuille |
triglochinine |
|
Trifolium repens |
feuille |
linamarine |
|
Sorghum bicolor |
feuille/graine |
dhurrine |
|
Taxus baccata |
feuille |
taxiphylline |
Parallèlement, d'autres composés de défense sont synthétisés uniquement lorsque la plante perçoit une attaque. Il s'agit généralement de molécules de faible poids moléculaire, toxiques pour les animaux ou les micro-organismes pathogènes. Ces composés portent le nom de phytoalexines. En dehors de ces molécules toxiques, les plantes synthétisent aussi des enzymes qui hydrolysent les parois des micro-organismes pathogènes.
La plupart des plantes sont capables de percevoir la présence d'un pathogène (maladie) ou un dommage causé à la plante (blessure) et d'induire, entre autres, la synthèse de phytoalexines. Généralement, cette perception se fait par l'intermédiaire de molécules spécifiques appelées éliciteurs. Elles peuvent être produites par le pathogène ou relâchées d'une paroi de cellule végétale endommagée.
Une cellule végétale attaquée peut aussi émettre un signal qui induira la synthèse de composés de défense dans le reste de la plante. Ainsi, lorsque le pathogène s'attaquera à la feuille suivante, celle-ci aura synthétisé des composés de défense et pourra mieux résister à l'attaque.
De plus, certaines plantes émettent des signaux volatils. Le méthyl-jasmonate est un signal volatil synthétisé par une plante blessée et, en concentration suffisante, il peut induire la synthèse de composés de défense chez une plante voisine.
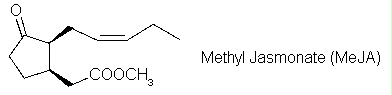
Pour pouvoir produire les phytoalexines, les plantes doivent induire la synthèse de plusieurs enzymes différentes. Une de ces enzymes est la polyphénol oxydase, qui apparaît en cas de blessure ou d'attaque par un pathogène. Cette enzyme provoque l'oxydation de plusieurs composés phénoliques présents dans la cellule végétale, ce qui les rend toxiques, en particulier pour les insectes herbivores.
(peut être modifié selon les indications de l'assistant)
Tous les groupes effectuent les expériences A et B.
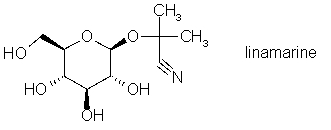
Le but de cette manipulation est de détecter la présence de glycosides cyanogéniques dans diverses espèces au moyen d'un test semi-quantitatif.
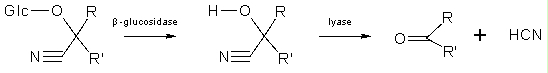
Nous utiliserons dans notre essais la linamarine (l’un des cyanogènes toxiques du manioc) :
Papier Test (préparation effectuée par l'assistant)
- Découper 5 rectangles de 8x12 cm de papier buvard.
- Sous la hotte, préparer 25 ml d'une solution de chloroforme à 1% P/V d'éthyle acétoacétate de cuivre.
- Sous la hotte, préparer 25 ml d'une solution de chloroforme à 1% P/ V de di-(4-diméthyl-aminophenyl)-méthane (! : produit toxique, port des gants et des lunettes obligatoire).
- Mélanger ces deux solutions dans une grande boîte de Pétri en verre.
- Tremper les rectangles de papier buvard dans le mélange, et les laisser sécher sous la hotte jusqu'à évaporation complète du solvant. (! : même sec ne manipuler ces papiers qu'avec des gants.)
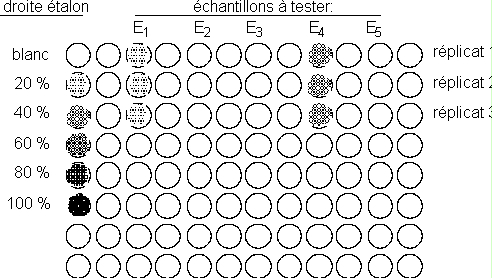
Gamme étalon
- Préparer dans 6 tubes Eppendorf, 100 µl de dilutions aqueuses à 0, 20, 40, 60, 80 et 100% de la solution stock de linamarine (concentration du stock: 2 mM)
2 échantillons différents E1, E2 sont fournis à chaque séance. Les étudiants sont invités à amener d'autres matériels à tester (feuilles, graines, fruits, ...)
- Découper les matériels aux dimensions des micro puits (Emporte-pièces, ciseaux, scalpel, ...).
- Peser chaque échantillon sur la balance de précision. Noter le poids. Utiliser entre 1 et 20 mg par puits
- Les déposer dans les puits de la micro plaque, sans oublier de noter la disposition des divers échantillons sur la micro plaque.
- Ajouter 100 µl d'eau distillée dans les puits contenant les échantillons.
- Écraser les échantillons avec une baguette de verre.
- Déposer 20 µl de linamarase (10 milliunités) dans les puits destinés à recevoir la gamme étalon.
- Déposer ensuite dans ces 6 puits les solutions de la gamme étalon
- Poser le papier test sur les puits, et le recouvrir d'une feuille de papier absorbant pliée en 4.
- Fermer le couvercle de la micro plaque et le maintenir au moyen d'élastiques.
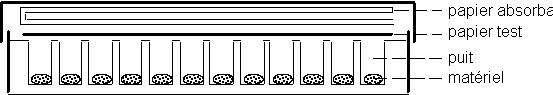
- Placer dans l'incubateur à 35°C pendant 1 heure.
Le réactif contenu dans le papier test réagit avec le cyanure dégagé par les échantillons et prend une coloration bleue. L'intensité de la coloration est proportionnelle à la quantité de cyanure dégagé, et peut être comparée à la gamme étalon. Une mole de linamarine dégage une mole de cyanure sous l'effet de la linamarase.
Pour chaque échantillon, estimer la quantité de cyanure dégagée, et la rapporter au poids frais de matériel.
Comment les insectes ou les pathogènes peuvent-ils réagir contre les glycosides cyanogéniques ?
Quels sont les autres types de défense utilisés par la plante?
Certaines plantes contiennent-elles suffisamment de glycosides cyanogéniques pour mettre en danger la santé d'un être humain? Calculer combien de grammes de votre échantillon le plus actif faudrait-il manger pour absorber la dose mortelle d'HCN, soit 70 mg?
Le but de cette expérience est de mettre en évidence l'induction de la polyphénol oxydase dans des plantules de tomate blessées (B), dans des plantules mises en présence de méthyl-jasmonate (C).
Traitement: Effectué 48h à l'avance par le groupe précédent ou par l'assistant.
Placer les plants de tomate dans les bocaux de conserve.
a) Blessure: écraser une des deux feuilles de la plantule avec une grande brucelle en travers des nervures. marquer la feuille blessée avec un fil.
b) méthyl-jasmonate: déposer 1 μl de méthyl-jasmonate 10% (w/v) sur un coton-tige et le placer dans le bocal sans qu'il touche les plantules.
Fermer les bocaux et les placer 48 h sous lumière constante.
Échantillons à observer:
A: feuille d'une plantule témoin
B: feuille blessée
C: feuille d'une plantule mise en présence de méthyl-jasmonate
- prélever une feuille pour chacun des différents traitements.
- broyer dans un tube eppendorf dans 200 μl de tampon P
- centrifuger 5 min.
- découper un fragment de membrane de nitrocellulose de la taille d'une lame de microscopie et le déposer sur la lame.
- déposer différentes quantités de surnageant (4 et 8 μl) sur la membrane.
(Les protéines solubles extraites lors du broyage vont se fixer sur la membrane de nitrocellulose, chargée positivement, par interactions électrostatiques.)
- rincer doucement la membrane dans le tampon P
- placer la lame dans une boîte de pétri
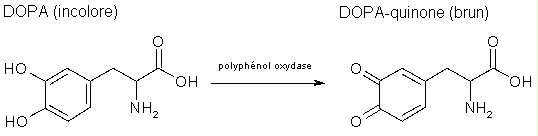
- recouvrir de la solution de DOPA (dihydroxyphényalanine)
- incuber la boîte de pétri ouverte 20 min à 26°C
- rincer la membrane à l'eau et garder humide pour l'observation.
Faire un tableau montrant la présence de la polyphénol oxydase dans les tomates ayant subi les différents traitements. Estimer la quantité relative présente dans chaque cas (Exprimer avec +, ++, etc.). Expliquer et commenter ces résultats.
- quel est l'avantage pour la plante d'induire des composés de défense dans les feuilles non attaquées?
- quel pourrait être l'avantage "d'avertir" les plantes voisines de la présence d'un pathogène?
- pouvez-vous imaginer une expérience qui permette un dosage quantitatif de la quantité de polyphénol oxydase induite lors de chaque traitement?
La photosynthèse est la conversion de l’énergie lumineuse en énergie chimique, dans les chloroplastes des cellules eucaryotes. L’absorption de cette énergie est réalisée, au niveau de la membrane des thylakoïdes, par l’intermédiaire de molécules sensibles à lumière: les chlorophylles.
La chlorophylle est localisée, à l'intérieur de la cellule végétale, dans les chloroplastes. Ces chloroplastes font partie des organites des cellules eucaryotes, ils n'existent que chez les végétaux au sens large. Ils sont issus de la transformation de proplastes sous l'effet de la lumière. Les proplastes, non soumis à la lumière, donnent des étioplastes.

Un chloroplaste est composé d'une double membrane isolant un milieu interne (le stroma) du contenu cellulaire. Un système membranaire, issu de la membrane interne, forme un ensemble de longs sacs aplatis, les thylakoïdes, baignant dans le stroma. L'empilement des thylakoïdes porte le nom de granum. Dans le stroma on trouve de l'ADN, des ribosomes, des globules lipidiques ainsi que de l'amidon. Les molécules de chlorophylles sont localisées sur la membrane des thylakoïdes.
Le chloroplaste proviendrait d'une relation symbiotique entre une cyanobactérie et une cellule eucaryote primitive.
Les pigments sont présents dans des structures protéiniques organisées en photosystèmes (PSI et PSII). Ils sont présents, en particulier dans une antenne collectrice, composée de chlorophylle "a" et "b" ainsi que de caroténoïdes et xanthophylles, et dans un centre réactionnel comprenant uniquement d'une paire de chlorophylle a.
La chlorophylle "a" existe en deux variantes selon les protéines qui lui sont associées : la P680 qui absorbe des longueurs d'ondes de 430 nm dans le bleu et de 680 nm dans le rouge (PSII) et la P700 qui absorbe à 700 nm au lieu de 680 nm (PSI). Les autres pigments accessoires ont un spectre d'absorption décalé : la chlorophylle "b" absorbe vers 460 nm et 600 nm, les caroténoïdes absorbent dans les orangés-jaunes et le bleu, les phycobillines dans le vert et le bleu.
Quant un pigment absorbe un photon d'énergie donnée (qui est fonction de la longueur d'onde), un électron change de niveau d'énergie. La molécule est alors dite excitée car l'électron ; elle se trouve à un niveau d'énergie supérieur très instable. Cet électron revient d'ailleurs rapidement à son état initial en 10-9 s . Se faisant, il libère son énergie sous forme de chaleur, et pour certains pigments comme la chlorophylle également sous forme d'un photon, ce qui entraîne un phénomène de fluorescence (le photon émis est d'énergie plus faible que celui qui a excité le pigment, donc de plus grande longueur d’onde),.
Dans le chloroplaste, l'électron instable peut être piégé par une molécule acceptrice.. Cette molécule, un accepteur d'électron, est située au côté du centre réactionnel dans les photosystèmes. Les pigments accessoires comme les caroténoïdes ne font que transmettre l'énergie reçue par les chlorophylles a et b de l'antenne collectrice, ils n'émettent pas d'électrons. Le rôle de l'antenne collectrice, possédant un spectre d'absorption large, est de récupérer le maximum d'énergie et de la focaliser vers le centre réactionnel.
Il existe deux types de photosystèmes : le PSI (riche en chlorophylle a P700) et le PSII (riche en P680). Leur composition en pigments est différente.
Au niveau de PSII va s'opérer une étape majeure de la photosynthèse : la photolyse de l'eau. À chaque fois que PSII est photo-oxydé, l'eau lui fournit un électron pour compenser la perte que le photosystème vient de subir et permettre sa régénération. En donnant un électron (e-) l'eau subit une réaction d'oxydation :
|
2H2O ----> 2OH-+ 2 H+ Les H+ vont s'accumuler dans l'espace intra-thylakoïdien |
L’eau est donc le donneur d’électron primaire de la photosynthèse et l'oxygène libéré à la suite de son oxydation n'est en fin de compte qu'un déchet de la photosynthèse.
A l'état excité PSII transmet un électron vers PSI par l'intermédiaire d'une suite de réactions d'oxydoréduction faisant intervenir des transporteurs d'électrons comme la plastoquinone, l'accepteur primaire d'électrons, les cytochromes et la plastocyanine. L'électron au cours de ces différents transferts perd un peu d'énergie. Cette énergie est utilisée par certains transporteurs pour amener des protons H+ du stroma vers l'espace intra-thylakoïdien. Quant cet électron arrive sur le PSI, il va permettre la régénération de la molécule de P700 photo-oxydée par la lumière. L'électron produit par le PSI va également suivre une chaîne de transporteurs d'électrons qui cette fois-ci fait intervenir un pigment caroténoïde (X) et une ferredoxine. L'électron est alors transmis à une NADP réductase qui va, comme son nom l'indique, réduire un accepteur d'électron, le NADP+ en NADPH + H+
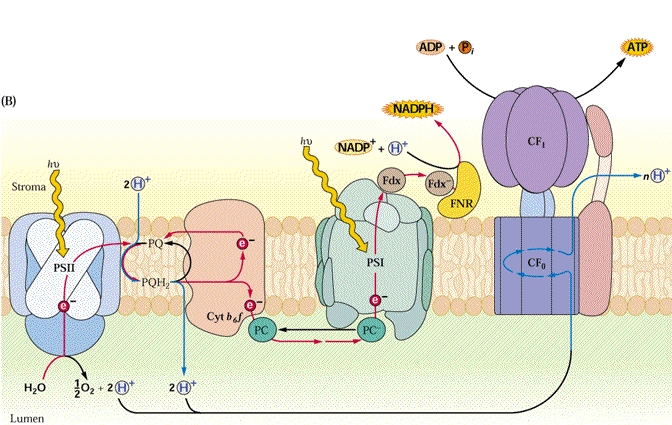
Figure 1 : Schéma de la phase lumineuse de la photosynthèse.
L’énergie lumineuse est captée par des complexes de chlorophylles (light harvesting complexes) et est transmise au cœur du photosystème II (PSII), à la chlorophylle P680. Cette chlorophylle énergisée arrache deux électrons à une molécule d’eau. Dans les conditions optimales, ces électrons vont réduire une molécule acceptrice, la phéophytine, puis être transférés grâce à une série de transporteurs (plastoquinone, cytochrome b6/f, plastocyanine) vers le photosystème I (PSI). Ce transfert est associé avec la création d’un gradient de proton et conduit à la production d’ATP, par couplage d’une ATPase à proton (photophosphorylation acyclique). Les light harvesting complexes transmettent aussi leur énergie au cœur du PSI, la chlorophylle P700. Cette énergie permet d’arracher deux électrons à la plastocyanine pour réduire la ferredoxine, qui est ensuite impliquée dans la réduction du NADP en NADPH2. Ce pouvoir réducteur (NADPH2) pourra être ensuite utiliser pour la réduction du CO2 et la production de sucres dans la phase sombre de la photosynthèse.
Le NADPH est une molécule transporteuse d’électron très présente dans de nombreuses réactions biologiques :
|
NADP+ + 2H+ + 2e- ----> NADPH + H+ |
Dans certains cas, tel un excès de NADPH + H+, les électrons peuvent suivre un trajet cyclique qui n'implique que le photosystème I.
La ferredoxine, au lieu de fournir les électrons à la NADP réductase, va les transmettre à la plastoquinone par l'intermédiaire d'un cytochrome. Les électrons suivent alors la première chaîne de transporteurs qui les fait revenir au photosystème I, où ils vont combler les vides qu'ils avaient laissés. Ce trajet cyclique permet d'accumuler des protons supplémentaires dans l'espace intra-thylakoïdien sans réduire de NADP+ mais en favorisant la production d’ATP
L'accumulation des protons H+ dans l'espace intra-thylakoïdien entraîne un déséquilibre avec la concentration des protons du stroma. Le pH de l'espace intra-thylakoïdien descend à 5 alors que celui du stroma monte à 8. . Le gradient de proton ainsi établi favorise toute réaction qui permet de l’annuler. C’est ainsi que le complexe de couplage (CF1 en forme de sphère pédonculée) est activé. Ce complexe ATPase forme un canal à proton au travers de la membrane thylakoïdale. Lors du passage de trois protons, un groupement phosphate est ajouté par le CF1 à de l'ADP (adénosine diphosphate), créant ainsi de l'ATP (Adénosine triphosphate).
La photosynthèse produit des équivalents réducteurs et de l'ATP.
Métabolisme des végétaux ; physiologie et biochimie – Gerhard Richter – PPUR, Lausanne, 1993 – ISBN 2-88074-231-5 – Voir en particulier le chapitre 3 intitulé « Autotrophie des organismes végétaux ».
Sur Internet : http://www1.biologie.uni-hamburg.de/b-online/e00/default.htm
On trouvera un guide de laboratoire en ligne à l’adresse : http://web.grinnell.edu/courses/bio/qubitmanual/ (ce guide est en anglais)
Groupes I.1 et II.1 : A & B
Groupes I.2 et II.2 : C
séparer par chromatographie et identifier par analyse spectrale les pigments présents dans une feuille et associés à la photosynthèse
Toute l’extraction sera effectuée sous hotte à aspiration (solvants toxiques et inflammables).
Couper 4 à 5 grammes de feuilles.
Broyer dans le mortier, sur glace, après avoir ajouté du sable de quartz et du carbonate de calcium (pour neutraliser l’acidité du milieu), jusqu’à obtention d’une pâte homogène.
Ajouter 10 ml d’acétone et continuer à broyer délicatement (éclaboussures) quelques minutes, jusqu’à obtention d’une solution vert-foncée.
Laisser un peu décanter et filtrer la solution dans une ampoule à décanter (Bien vérifier que le robinet est fermé).
Ajouter un volume d’éther de pétrole.
Boucher l’ampoule, la retourner en tenant le bouchon et agiter par petites rotations après avoir ouvert le robinet.
Agiter ainsi jusqu’à ce que le surnageant (l’éther de pétrole) soit devenu vert-foncé.
Fermer le robinet, reposer l’ampoule sur son support et ajouter quelques ml d’eau distillée. Cette opération a pour but de faire repasser les pigments caroténoïdes et xanthophylles dans la phase éther-pétrole.
Agiter de nouveau comme précédemment (sans oublier de dégazer et en tenant bien le bouchon !)
Remettre l’ampoule sur son support et attendre que les deux phases soient bien séparées.
Enlever le bouchon de l’ampoule et éliminer la phase inférieure.
Effectuer encore deux lavages avec un volume égal d’eau distillée.
Récupérer la phase éther-pétrole dans un tube propre (éviter absolument toute trace d’eau)
Faire évaporer l’éther de pétrole sous un courrant d’air sous pression.
Reprendre le culot dans 1 ml d’éther de pétrole.
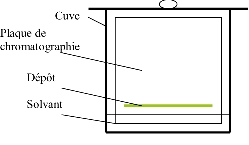
Schéma de l’installation (figure) :
La chromatographie sera effectuée dans une cuve préalablement équilibrée dans le tampon suivant :
Ether de pétrole : 3 volumes
Acétone : 1 volume
Chloroforme : 1 volume
Prendre une plaque de silice au four.
Tracer un trait au crayon à papier, à 1 cm du bord, sans gratter la couche de silice.
Déposer sur la plaque environ 200 μl de la phase éthéro-pétrolique obtenue précédemment, sur le trait tracé, en essayant de faire un dépot le plus fin possible. Le dépôt peut être effectué en plusieurs fois.
Laisser sécher à l’air quelques minutes.
Placer la plaque de silice dans la cuve, en vérifiant bien que le dépôt ne trempe pas dans le solvant.
Attendre que le front de migration soit parvenu au somment de la plaque.
Noter immédiatement la distance entre le dépôt et le front de migration.
Noter les distances de migration des différents pigments visibles.
Sur la plaque, racler la silice au niveau de chaque bande de pigment. Récupérer ainsi chaque bande indépendamment dans un tube propre et ajouter 1 ml d’acétone.
Effectuer le spectre d’aborption de chaque pigment entre 400 nm et 750 nm.
Etudier le fonctionnement des chloroplastes isolés, leur teneur en pigments photo actifs et, enfin, vérifier qu’ils sont bien capables de réduire un accepteur d’électron bien choisi tout en produisant de l’oxygène selon une réaction stœchiométrique
Prélever 5 g de feuilles fraîches. Les laver, éliminer les nervures principales et le pétiole. Les broyer dans un mortier maintenu au froid en présence de 10 ml de milieu d'extraction (M) dont la composition est la suivante: tampon Tricine 30 mM, pH 8, EDTA 1 mM, MgCl2 1 mM, NaCl 10 mM, saccharose 400 mM. Filtrer le broyat sur de la gaze et centrifuger le filtrat à 2000 g pendant 10 minutes. Éliminer délicatement le surnageant et reprendre le culot de chloroplastes dans 10 ml de milieu M ; les remettre en suspension à l'aide d'un pinceau, les conserver au frais à l'obscurité.
Prélever 10 µl de la suspension de chloroplastes, les mettre en solution dans 1 ml d'acétone à 80 %. Décanter et doser les chlorophylles totales. Pour ce dosage, effectuer le 0 de densité optique avec de l'acétone à 80 % et mesurer la D.O. de l'extrait à 645 nm, 663 nm et 652 nm.
Calculer la teneur en chlorophylles totales de la suspension en utilisant la formule de ARNON (1949). Tous les résultats obtenus seront basés sur la teneur en chlorophylle totale des extraits.
Formule de ARNON :
chl a = (0,0127 x D.O. 663) - (0,00269 x D.O. 645) mg.ml-1
chl b = (0,0229 x D.O. 645) - (0,00468 x D.O. 663) mg.ml-1
chl totale = (0,0202 x D.O. 645) + (0,00802 x D.O. 663) mg.ml-1
|
ou chl totale = |
mg.ml-1 |
|
|
34,5 |
Les résultats sont exprimés en mg.ml-1 de chlorophylle. Tenir compte de la dilution initiale (10 µl dans 1 ml).
Cette réaction, qui résume en fait une série de processus, peut se traduire par l'équation suivante :
H2O + A + lumière + chloroplastes + 2 e- ------> H2A + 1/2 O2
« A » qui est un accepteur d'hydrogène est donc un oxydant.
Au cours de cette manipulation, on utilisera comme réactif de Hill le 2,6 DCPIP (2,6 dichlorophénolindophénol) qui est bleu sous forme oxydée et incolore sous forme réduite. La courbe de réduction du colorant est obtenue en mesurant en fonction du temps la diminution de D.O. d'une solution de 2,6 DCPIP à 600 nm.
|
|
||
|
forme quinonique oxydée colorée |
forme réduite incolore leucodérivé |
|
Le NADP+ est l'oxydant de Hill naturel. Le réactif de Hill le plus utilisé in vitro est le 2,6 DCPIP. Sa forme oxydée, bleue, présente une bande d'absorption dans le rouge, qui disparaît quand il est réduit et il passe sous forme incolore = leucodérivé.
Étant donné que le colorant peut être réduit par des processus étrangers à l'activité photosynthétique des chloroplastes, on vérifiera la présence éventuelle de réactions de réduction du 2,6 DCPIP autres que la réduction photochimique en étudiant, en présence de chloroplastes, la réduction du colorant dans des tubes maintenus à l'obscurité.
Préparer successivement 4 tubes expérimentaux comme suit.
Verser dans l'ordre et respecter les délais indiqués.
|
tube 1 : |
- 2,6 ml de tampon |
|
- quelques cristaux de Na2S2O4 |
|
|
- 0,2 ml de la suspension de chloroplastes |
Suivre la densité optique à 600 nm. Régler le 0 avec ce premier tube. Ce réglage est valable pour toute la durée de l'expérience.
|
tube 2 : |
- 2,6 ml de tampon |
|
- 0,2 ml de 2,6 DCPIP 5.10-4 M |
|
|
- 0,2 ml de suspension de chloroplastes |
Agiter et insérer immédiatement le tube dans le spectrophotomètre. Lire la densité optique (D.O.). Placer ensuite ce tube à la lumière et effectuer des mesures de D.O,. toutes les minutes pendant 15 à 20 minutes.
|
tube 3 : |
- 2,6 ml de tampon |
|
- 0,2 ml de 2,6 DCPIP 5.10-4 M |
|
|
- 0,2 ml de suspension de chloroplastes |
Agiter et insérer dans le spectrophotomètre et lire aussitôt la D.O. Placer ensuite rapidement le tube à l'obscurité. Effectuer des mesures de D.O. toutes les 4 minutes.
Au bout de 12 minutes, effectuer une lecture et placer le tube à la lumière. Suivre les variations de D.O. toutes les 2 minutes pendant 10 minutes environ.
|
tube 4 : |
- 2,2 ml de tampon |
|
- 0,2 ml de 2,6 DCPIP 5.10-4 M |
|
|
- 0,4 ml de DCMU 0.5 mM (dichlorométhylurée) |
|
|
- 0,2 ml de suspension de chloroplastes |
Agiter et placer ce tube à l'obscurité pendant environ 10 minutes. Puis, ajouter 0,2 ml de 2,6 DCPIP. Agiter et lire aussitôt la D.O. Placer ensuite ce tube à la lumière. Suivre, pendant 15 minutes environ, les variations de D.O. toutes les 2 minutes. Conclure.
Présenter les résultats obtenus sous forme de tableau. Tracer les courbes de variation de D.O. pour les tubes 2, 3 et 4 en fonction du temps.
- Calcul de la quantité d'oxygène dégagé :
La réduction du 2,6 DCPIP due réellement à la réaction de Hill correspond à la valeur corrigée de la variation de D.O. à la lumière pour une période de temps t0 à t1 soit:
Δ D.O.lum = D.O.t0 - D.O.t1
par la variation de D.O. observée à l'obscurité pendant la même période, soit:
Δ D.O.obsc. = D.O.'t0 - D.O.'t1
ce qui correspond à
ΔD.O.lum -Δ D.O.obsc. = Δ D.O. due réellement à la réaction de Hill,
La quantité de 2,6 DCPIP réduite exprimée en mole sera :
![]()
où X représente le volume réactionnel- en ml.
d = épaisseur de liquide que traverse le faisceau ou trajet optique (l cm).
ε = coefficient d'extinction moléculaire de 2,6 DCPIP à 600 nm, soit 1,26.107.cm-1.mole-1.ml.
On exprimera les résultats en volume d'oxygène dégagé, par heure, soit :
![]()
et par gramme de substance fraîche :
![]()
1/0,4 représente la quantité de chloroplastes prélevée pour l'expérience et ramenée à 1 ml et Pg/Vs représente le rapport du volume de suspension recueillie au poids de substance fraîche pesée. On peut exprimer, les résultats en mmoles d'O2 dégagé h-1 mg-1 de chlorophylle en se rapportant au dosage préalablement effectué.
Etudier le devenir de l’énergie lumineuse absorbée par les systèmes photosynthétique et estimer le bon fonctionnement de la photosynthèse (taux de transport des électrons) ; ce dernier paramètre est un élément important qui caractérise l’efficacité des plantes dans leur milieu (études écologiques, optimisation des rendements agricoles)
Quand un photon excite une molécule de chlorophylle, un électron est porté à un niveau d’énergie supérieur. Cette énergie peut être dissipée sous forme de chaleur (D), utilisée pour des réactions photochimiques (P) ou il peut être émise, sous forme d’un nouveau photon, par fluorescence (F). P est le quotien photosynthétique et F, le taux de fluorescence.
Pour un photon on fixe, arbitrairement, la somme des énergies à 1 (un Einstein) :
P + F + D = 1
On se trouve en condition saturante de lumière, la presque totalité de l’énergie lumineuse est dissipée sous forme de fluorescence et de chaleur. On a donc la formule suivante :
Fmax + Dmax = 1
De plus, expérimentalement, il a été montré que le rapport entre l’énergie libérée sous forme de chaleur et l’énergie libérée par fluorescence était relativement constant, quelque soit l’intensité lumineuse. Ce qui nous donne l’équation suivante :
Dmax/Fmax = D/F
Dmax = 1 – Fmax donc D/F = (1 – Fmax)/Fmax puis D = F(1 – Fmax)/Fmax
Et si on remplace la valeur de D dans la première équation on obtient :
P + [F(1 – Fmax)/Fmax] + F = 1 puis P = 1 – F – F(1-Fmax)/Fmax
Et si on ramène tout au même dénominateur :
P = (Fmax – F*Fmax – F + F*Fmax)/Fmax
Et après simplification, l’énergie utilisée pour les réactions photochimiques est :
P = (Fmax – F)/Fmax
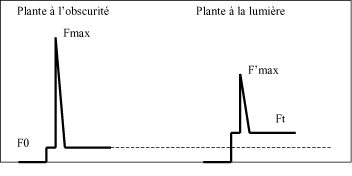
Le quotien P peut être utilisé pour calculer le taux de transport d’électrons. Etant donné qu’un photon permet l’excitation d’un électron dans la chlorophylle, P représente la part de l’énergie utilisée pour le transport des électrons lors de la photosynthèse. Le taux de transfet des électrons (TRE) est fonction du flux de photons photosynthétiquement actif (PAR) arrivant à la surface de la feuille. De plus, 84% de la lumière incidente à la surface de la feuille peut être absorbée par la cholorophylle et 50% des photons activent les molécules de chlorophylle du PS II et 50%, celles du PS I.
Ainsi on peut avoir une estimation du TER par la formule suivante :
TRE = P x PAR x 0.42
Le PAR étant mesuré par un photosenseurr lors de l’expérimentation.
Les deux groupes effectueront les mêmes expériences mais avec des matériels végétaux différents.
Les plantes seront maintenues à l’obscurité avant le début de l’expérimentation.
Ouvrir le fichier « fluores.mbl »
Régler le zéro du fluoromètre : allumer la lumière et collecter les données, à vide.
Prendre une feuille d’une plante gardée à l’obscurité et la placer dans la chambre et entourer la chambre d’aluminium pour garder la feuille à l’obscurité.
Mettre en marche le boitier de contrôle de la lumière, mais ne pas allumer la lampe à incandescence. Allumer la lampe LED.
Lancer l’enregistrement des mesures en cliquant sur « collect ».
Régler l’amplificateur du fluoromètre pour lire le signal entre 0,2 et 0,6.
Noter la valeur F0. Déclencher un flash (5000 μmol quanta/m2/s) et noter la valeur Fmax.
Allumer la lampe à incandescence et régler l’intensité lumineuse à environ 200 μmol quanta/m2/s. La fluorescence augmente jusqu’à une valeur proche de Fmax. A ce moment, déclancher un flash pour obtenir la valeur F’max.
Appliquer ensuite des flashs à répétition en réglant les pulses sur automatique. Quand le signal de fluorescence atteind une valeur stable arrêter l’enregistrement et sauver les données.
L'oxygène oxyde les métabolites en produisant de l'énergie, un peu comme un feu produit de la chaleur en consommant l'oxygène et en dégradant de la matière organique. Au niveau cellulaire, l'énergie issue de la respiration n'est que partiellement transformée en chaleur, elle est principalement convertie en énergie chimique sous forme d'ATP (adénosine triphosphate) lors de la phosphorylation oxydative.
L’organite responsable dans la respiration cellulaire est la mitochondrie. Outre la production et la conversion d'énergie, la respiration, en produisant de l’eau à partir de l’oxygène permet à la cellule de se protéger contre ce dernier qui est un élément très toxique.
C'est un organite en forme de bâtonnet de 7 µm de long et 1 µm de diamètre (il ressemble à un bacille). Sa distribution dans la cellule est généralement homogène. La mitochondrie possède deux membranes séparées par un espace inter membranaire. La membrane interne est fortement plissée vers l'intérieur de la mitochondrie, et forme les crêtes mitochondriales : ces replis augmentent considérablement la surface de cette membrane. Ces enveloppes délimitent une matrice mitochondriale. On y trouve un ADN mitochondrial, des ribosomes spécifiques ainsi que divers granules.

Du nombre de crêtes dépend l'activité respiratoire. En effet c'est au niveau de la membrane interne que vont s'opérer les diverses réactions de la chaîne d'oxydation. Cette membrane porte d'ailleurs des complexes enzymatiques en forme de sphères pédonculées (dirigées vers la matrice). Ce sont des ATP synthases ou ATPases. Ce complexe enzymatique (ou complexe F0-FI) peut, suivant les circonstances, synthétiser ou dégrader l'ATP (on le retrouve dans les thylakoïdes des chloroplastes).
La membrane externe comporte plusieurs protéines membranaires :
des protéines canaux qui permettent l'entrée de petites molécules
des transporteurs
des kinases, consommatrices d'ATP
des phospholipases, des phosphatases
des ATP-acétylCoA synthases
des NADH réductases
La membrane interne possède :
des transporteurs spécifiques. La membrane interne est imperméable (sauf aux lipides). Il existe donc un transporteur spécifique pour chaque molécule devant traverser cette membrane (même l'ADP et l'ATP).
les enzymes permettant la formation d'ATP :
Enzymes de la chaîne respiratoire
Enzymes de la ß-oxydation des lipides
l'ATPase
La matrice contient les enzymes nécessaires au bon fonctionnement de la synthèse de protéines mitochondriales ainsi que les enzymes du cycle de Krebs (cycle des acides tricarboxylique) et de la ß-oxydation des lipides.
Pour que la synthèse d'ATP puisse avoir lieu, la mitochondrie doit générer un grand nombre de donneurs d'électrons ou agents réducteurs. Ces donneurs sont formés (ou plutôt "rechargés") au cours de la dégradation de divers métabolites en acétyl-CoA puis au cours de la dégradation de cet acétyl-CoA dans le cycle de Krebs.
Le carbone des métabolites (glucose, acides aminés, acides gras ainsi que lactate et pyruvate) se retrouve dans le groupe acétyle (2C) de l’acétylcoenzyme A. La formation de l’acétyl-CoA à lieu dans le cytoplasme et génère des équivalents réducteurs sous forme de donneurs d'électrons (NADH+H+, et NADPH+H+) qui seront importés dans les mitochondries
Les réactions peuvent suivre deux voies métaboliques différentes qui ont lieu dans le cytoplasme :
La glycolyse : Le glucose est phosphorylé puis transformé en glycéraldéhyde-3P et dihydroxyacétone-P (deux isomères en équilibre). Ces réactions consomment 2 molécules d'ATP. Le glycéraldéhyde-3P est ensuite transformé en phosphoénol-pyruvate puis en pyruvate. Cette suite de réaction produit 2 molécules d'ATP et la réduction d'un donneur d'électron (NAD+). Pour une molécule de glucose (2 x 3C), ce sont donc 4 ATP qui sont formés. La glycolyse d’une molécule de glucose produit donc 2 ATP, 2 NADH+H+.
La voie des pentoses phosphates : Le glucose subit une suite de réactions permettant la réduction de deux NAD+ en NADH+H+. Ces réactions aboutissent à la formation de Fructose-6P, qui va s'intégrer dans la voie de la glycolyse (c'est le premier produit de transformation du glucose-6P dans cette voie). L'association voie des pentoses et glycolyse permet ainsi de former 2 NADH+H+ supplémentaires.
Le pyruvate formé est ensuite transféré dans la matrice mitochondriale où il est transformé en acétyl-CoA par perte de CO2 (la première molécule de gaz carbonique produite lors de la respiration) et addition d'un groupement CoA sous l'action du complexe enzymatique de la pyruvate déshydrogénase (Pyruvate décarboxylase (E1) + dihydrolipoyl transacétylase (E2) et dihydrolipoyl déshydrogénase (E3)).
L'acétyl-CoA va être condensé avec l'acide oxaloacétique pour former de l'acide citrique. Au cours de cette réaction, il perd son groupement CoA. Une série de réactions d'oxydation va permettre de reformer de l'acide oxaloacétique mais surtout de former 4 nouveaux donneurs d'électrons (3 NADH+H+ et 1 FADH2) et un ATP. Il y a donc 8 électrons libérés à chaque cycle (chaque donneur d'électrons en reçoit deux). Il y a également dégagement de deux molécules de CO2 (correspondant aux atomes de carbone de l'acétyl-CoA).
Les électrons provenant du cycle de Krebs vont être cédés par les donneurs d'électrons au niveau de la membrane interne de la mitochondrie. A ce niveau existe un complexe de molécules, constitué d'une série de transporteurs d'électrons. La plupart de ces transporteurs sont des protéines possédant un groupement non protéique sensible aux électrons (il est facilement oxydé ou réduit). Les électrons vont être transférés de transporteur à transporteur en libérant un peu d'énergie, à la fin ils vont servir à réduire l'oxygène :
Une flavoprotéine (FMN) est le premier accepteur d'électrons. Une fois réduite, elle les transfère à une protéine fer-soufre et retrouve son état oxydé. Une ubiquinone permet de transférer les électrons à un premier complexe de cytochromes. Puis, via le cytochrome c, les électrons sont transmis à un dernier complexe de transporteurs (le cyanure bloque une de ces étapes). La dernière enzyme de cette chaîne est une cytochrome c oxydase (cytochrome a). L'énergie perdue par les électrons à chaque transfert sur un autre transporteur, permet l'éjection de protons (H+) de la matrice vers l'espace intermembranaire de la mitochondrie.
La cytochrome-oxydase réduit l'oxygène (O2) en superoxyde (O2-). Ce superoxyde, si la stœchiométrie des électrons n’est pas correcte (4e-) peut produire du HO2 . Les superoxydes donnent naissance, dans le cytoplasme, à du H2O2 sous l’action d’une enzyme cuprique, la superoxyde dismutase. H2O2 est détruite par une catalase qui le transforment en H2O. Cette réaction libère de l'oxygène. Pour 2 molécules d'eau oxygénée (H2O2) il y a ainsi formation de deux molécules d'eau (H2O) et d'une molécule d'oxygène (O2).
Pendant ce temps les protons s'accumulent dans l'espace inter membranaire. Cette accumulation de charges positives provoque une différence de potentiel électrique entre l'espace inter membranaire et la matrice, ainsi qu'un gradient de pH. L'ATPase membranaire est ainsi activée et pour environ 3 protons transportés dans le gradient, on obtient la phosphorylation d'une molécule d'ADP en ATP. L'oxydation d'une seule molécule de NADH+H+ (ou FADH2) permet ainsi la formation de 3 ATP.
La dégradation d’une molécule de glucose aboutit, par la voie de la glycolyse à la formation de 38 ATP. Ce bilan n'est en fait pas fixe mais dépend d'un grand nombre de facteurs qui peuvent varier dans la cellule. Par exemple, certains organismes sont capables de contrôler la perméabilité de la membrane interne mitochondriale aux protons. En plus de l'ATPase, la membrane possède d’autres types de transporteurs de protons. Ceux-ci ne passent alors plus par l'ATPase et l'énergie libérée au cours du transport des électrons, est dissipée sous forme de chaleur. C’est ainsi que certains organes végétaux (comme les inflorescences des aracées) produisent de la chaleur et constituent des tissus thermogènes. L'ATP, ainsi que le citrate, intermédiaire du cycle de Krebs, exercent une rétro-inhibition sur la glycolyse. Cet effet permet de réguler la respiration.
Les mitochondries végétales possèdent en outre une voie d’oxydation alternative qui est résistante au cyanure (un inhibiteur de la cytochrome oxydase)
Dans certains cas, la cellule ne dispose plus suffisamment d'oxygène, elle est dans des conditions d'anaérobiose. C'est le cas, chez l’homme, lors d'effort de longue durée ; chez les plantes comme le riz, ceci peut se produire lors des phases de croissance immergées. La production d'ATP par la chaîne respiratoire devient alors impossible. La cellule possède toutefois le matériel enzymatique nécessaire pour en fabriquer (dans une moindre mesure) par fermentation. Cette voie est la seule possible pour les organismes anaérobies (beaucoup de bactéries).
La fermentation permet de terminer la glycolyse en régénérant les agents oxydants (NAD+ depuis le NADH+H+), non pas par la chaîne de transport d'électrons (inutilisable en l'absence d'oxygène), mais en réduisant le pyruvate en éthanol pour la fermentation alcoolique et, en lactate pour la fermentation lactique.
Métabolisme des végétaux ; physiologie et biochimie – Gerhard Richter – PPUR, Lausanne, 1993 – ISBN 2-88074-231-5 – Voir en particulier le chapitre 5 intitulé « Oxydations biologiques et gains d’énergie ».
Sur le web : http://www1.biologie.uni-hamburg.de/b-online/e00/default.htm
On trouvera un guide de laboratoire en ligne à l’adresse : http://web.grinnell.edu/courses/bio/qubitmanual/ (ce guide est en anglais)
Groupes I.1 et II.1 : A
Groupes I.2 et II.2 : B
Montrer que les plantes respirent ; mesurer les échanges de gaz (CO2 et O2) dans un organe non-photosynthétique la racine, et vérifier que la quantité d’oxygène absorbé par la respiration correspond à la quantité de CO2 dégagé
sectionner environ 80 fragments de racines de maïs, de 1 cm de long, prélevés préférentiellement dans la zone apicale.
Déterminer le poids frais de 4 lots d’environ 20 fragments
Placer les lots dans les coupelles avec du tampon
sac rempli d’air
pompe
régulateur de flux
chambre d’échanges gazeux
un tube à condensation
une colonne dessicative
un senseur à O2
un senseur à CO2
Ouvrir le fichier « CO2O2TP »
Calibrer le senseur à CO2 entre 0 et 500 ppm.
Aller dans le menu « calibration » et cliquer sur le port correspondant à la sonde à CO2.
Effectuer le Zéro en montant la pompe à air, suivi de la colonne piégeant le CO2 puis de la colonne dessicative.
Attendre la stabilisation et cliquer sur « Ok ». Régler ensuite la sonde pour une valeur connue : enlever la colonne piégeant le CO2 et fixer un sac d’air, contenant une quantité connue de CO2 (500 ppm) à l’entrée de la pompe à air. Attendre la stabilisation et cliquer sur « Ok ».
Régler le flux de gaz à 400 ml/mn
Lancer l’enregistrement.
Suivre les valeurs du CO2, de l’O2 et de la température jusqu’à obtention de valeurs stables.
Placer un lot pesé de racines dans la chambre d’échange gazeux. Prendre soin à ce qu’il y ait assez de tampon pour imbiber les fragments, sans pour autant les noyer complètement.
Attendre la stabilisation des valeurs de CO2 et de O2.
Placer ensuite la chambre d’échange gazeux sur la glace, et suivre les variations de production de CO2 en fonction de la température (jusqu’à une valeur de 10°C)
Chauffer ensuite la chambre avec un sèche-cheveux, suivre la variation de la production de CO2 en fonction de la température (jusqu’à une valeur de 40°C)
Attendre de nouveau la stabilisation de la température et de la production de CO2.
Ouvrir rapidement la chambre, et ajouter 1 ml de solution de NaF 0,5M. attendre la stabilisation de la production de CO2 et arrêter l’enregistrement.
Placer un nouveau lot de racine dans la chambre.
Lancer un nouvel enregistrement
Suivre la production de CO2 pendant quelques minutes dans les conditions standards.
Ouvrir rapidement la chambre et ajouter 1 ml de solution de glucose 1%.
Suivre la production de CO2 et attendre la stabilisation.
Augmenter la température de la chambre jusqu’à 40°C, grâce au sèche-cheveux et suivre la production de CO2.
Laisser la température revenir à sa valeur initiale. Ouvrir rapidement la chambre et ajouter 1 ml de solution de KCN 1mM (demander à l’assistant)
Suivre la production de CO2 jusqu’à stabilisation.
Démontrer la présence de l’enzyme de l’étape finale de la respiration, la cytochrome-oxydase et mesurer son activité dans des inflorescences de choux-fleur (aussi un organe non photosynthétique).
Peser rapidement 5g de matériel végétal (chou-fleur)
Couper rapidement en petits fragments à la l’aide d’une lame de scalpel.
Broyer, sur la glace, dans un mortier, en présence de 5 ml de tampon et d’un peu de sable de quartz
Filtrer le broyat et centrifuger 5min à 3000 rpm (élimination des parois cellulaires, des noyaux et des amyloplastes)
Reprendre le surnageant dans un nouveau tube et centrifuger 20 min à 8000 rpm
Reprendre le culot, enrichi en mitochondries, dans 1 ml de tampon de mesure.
Conserver l’extrait sur glace
Préparer 5 cuves pour le dosage spectrophotométrique de la manière suivante :
|
Tampon |
Triton X100 0.3% |
Cytochrome c réduit 0.3M |
Suspension de mitochondries |
|
|
Tube 1 |
790 μl |
100 μl |
100 μl |
10 μl |
|
Tube 2 |
780 μl |
100 μl |
100 μl |
20 μl |
|
Tube 3 |
770 μl |
100 μl |
100 μl |
30 μl |
|
Tube 4 |
760 μl |
100 μl |
100 μl |
40 μl |
|
Tube 5 |
750 μl |
100 μl |
100 μl |
50 μl |
Ajouter la suspension de mitochondries au temps t=0 et noter l’absorption à 550 nm toutes les minutes pendant 10 minutes
Effectuer la même série de test, sans mettre de détergent (Triton X100). Le remplacer par 100 μl de tampon.
En comparant ces nouvelles valeurs avec les précédentes, vous pourrez calculer un pourcentage de mitochondries intactes. En effet, le cytochrome c réduit ne peut pas pénétrer dans les mitochondries intactes et la cytochrome c oxydoréductase se trouve sur la face matricielle de la membrane interne des mitochondries. L’utilisation d’un détergent permet de mettre en contact l’enzyme avec le substrat. Sans détergent, seules les mitochondries abîmées présenteront une activité cytochrome c oxydase.
Exprimer les valeurs en unité d’absorbance/min
Sachant que 1 unité DO 550nm = 18.5 nmoles de cytochrome c, exprimer ensuite les valeurs en nmoles de cytochrome c oxydé/min/g de poids frais.
La source principale d'énergie sur la terre est le soleil. L'énergie du soleil est absorbée par la biosphère par un processus connu sous le nom de photosynthèse, qui se produit dans les plantes, les algues et quelques types de bactéries. La photosynthèse peut être définie comme le processus physico-chimique par lequel les organismes photosynthétiques utilisent l'énergie lumineuse pour synthétiser des composés organiques. Chez les plantes, les algues et les cyanobactéries, le résultat du processus photosynthétique est la fixation du dioxyde de carbone atmosphérique, sous forme de sucres, et le dégagement de l'oxygène moléculaire (photosynthèse oxygénique). Certains types de bactéries sont capables d’utiliser l'énergie lumineuse pour créer des composés organiques mais sans produire d'oxygène (photosynthèse anoxygénique). La photosynthèse fournit l'énergie et le carbone réduit exigés pour la survie de pratiquement toute la vie sur notre planète, aussi bien que l'oxygène moléculaire nécessaire à la vie aérobie. En outre, les combustibles fossiles actuellement brûlés pour fournir de l'énergie ont été produits par des organismes photosynthétiques il y a plusieurs dizaines de millions d’années.
Le processus photosynthétique a un impact profond sur l'atmosphère et le climat de la terre : il est à l’origine de la production d’une atmosphère oxydative et des profondes modifications des êtres vivant qui a accompagné l’augmentation de la teneur en oxygène. Tous les ans plus de 10% de tout le dioxyde de carbone atmosphérique est réduit en matière organique par les organismes photosynthétiques. On estime que les ceux-ci prélèvent 100 x 1015 grammes de carbone (C) par an (Houghton et Woodwell, 1989). C'est équivalent à 4 x 1018 kJ d'énergie libre stockée en carbone réduit, soit approximativement 0,1% de l'énergie incidente sur la terre par an. Le carbone réduit photosynthétiquement est principalement stocké sous forme de sucres, pour être ensuite oxydé par les organismes vivants pour leur survie (nutrition hétérotrophe), ou par la combustion. Actuellement, il y a plus de CO2 est déchargé dans l'atmosphère qu'il n’en est fixé par la photosynthèse, en raison de l’utilisation des stocks fossiles. Des mesures directes prouvent que tous les ans le contenu atmosphérique de carbone augmente d’environ 3 x 1015 grammes. Au cours des deux cents dernières années, le CO2 dans l'atmosphère a augmenté d'environ 280 parts par million (ppm) à son niveau courant de 360 ppm (0.0360%). Certains modèles prédisent que la quantité de CO2 dans l'atmosphère atteindra 700 ppm (0.0700%) dans le siècle courant. Les conséquences de ce changement rapide de notre atmosphère sont inconnues. Puisque le CO2 agit en tant que gaz à effet de serre, quelques modèles de climat prévoient que la température moyenne de l'atmosphère de la terre augmente de plusieurs degrés Celsius. Une telle augmentation de la température pourrait conduire à des changements climatiques drastiques.
J.B. van Helmont (1577 - 1644) montra qu’une plante cultivée dans un container pendant plusieurs années, gagnait bien plus de poids qu’il en avait été perdu dans son substrat. Le Britannique Hales (1677 - 1761) postula que l'air et la lumière étaient nécessaires pour la nutrition des plantes vertes, bien avant que la nature et la teneur des différents gaz de l’atmosphère ne fussent connues.
En 1771, Joseph Priestley, un ecclésiastique et chimiste anglais, montra que les plantes libéraient un gaz qui permettait la combustion et la respiration, ceci en brûlant une bougie sous une cloche de verre jusqu'à ce que la flamme soit éteinte. Lorsque l’on place une plante sous la cloche de verre, la combustion de la bougie est entretenue. Priestley effectua ensuite la même expérience avec une souris placée sous la cloche avec une plante : il déduisit que les plantes « purifiaient » l’air. Bien que Priestley n'ait pas eu connaissance de l'oxygène moléculaire, son travail a prouvé que les plantes rejètent de l'oxygène dans l'atmosphère. S’inspirant du travail de Priestley, Jan Ingenhousz, un médecin hollandais, démontra que la lumière du soleil était nécessaire pour la photosynthèse et que seules les parties vertes des plantes pouvaient libérer l'oxygène. Pendant cette période Jean Senebier, botaniste et naturaliste suisse, découvrit que le CO2 était nécessaire pour la croissance photosynthétique et Nicolas-Théodore de Saussure, chimiste et physiologiste suisse, prouva que l'eau participait à la production photosynthétique de matière.
En 1845, Jules Robert von Mayer, médecin et physicien allemand, postula que les organismes photosynthétiques convertissaient l'énergie lumineuse en énergie chimique libre.
En 1905, le physiologiste anglais FF. Blackman démontra que la photosynthèse était un processus en deux étapes : la photolyse de l’eau qui conduit à la production d’oxygène est dépendante de la lumière (phase claire), tandis que la fixation du CO2 peut se produire à l’obscurité et est dépendante de la température (phase obscure).
L'énergie produite lors de la phase claire (voir chapitre sur la chlorophylle) va permettre à des réactions de synthèse de molécules organiques d'avoir lieu. Ces réactions constituent le cycle de Calvin, nommé ainsi en l’honneur de son découvreur, Melvin Calvin, prix Nobel de chimie 1961. On parle de phase obscure pour indiquer que ces réactions n'ont pas besoin de lumière pour être effectuée. Il ne faut pas en déduire que la lumière n’influence pas ces mêmes réactions.
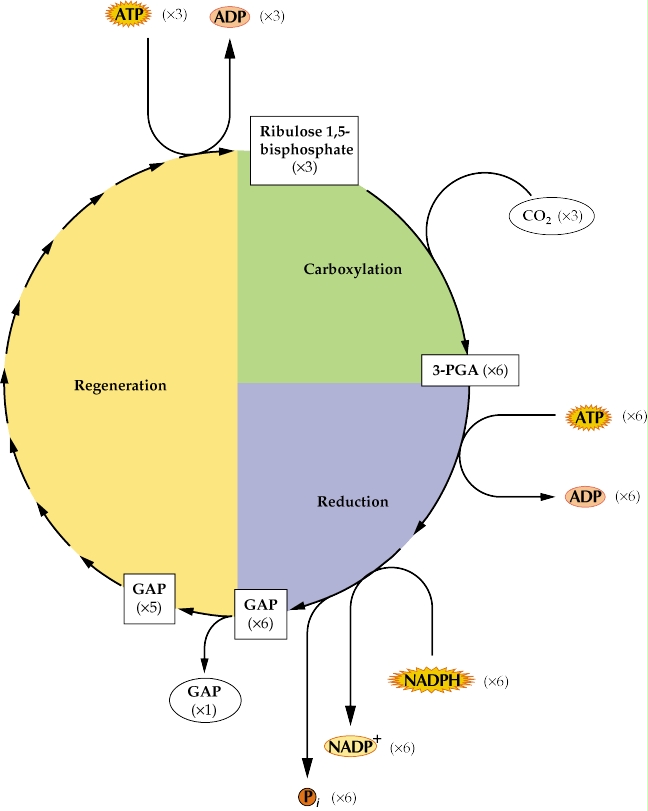
Grâce à la RUBISCO (Ribulose bisphosphate Carboxylase/Oxygénase), le CO2 atmosphérique va s'associer à une molécule à cinq carbones (C5), le ribulose diphosphate ou RuDP et produire deux molécules d’acide phosphoglycérique (PGA), molécules à trois carbones (C3) : c’est la fonction carboxylase de la RUBISCO. Ces PGA sont ensuite transformés en trioses phosphates (phosphoglycéraldéhyde ou GAP) par addition d'un groupement phosphate provenant d'une molécule d'ATP et réduction par une molécule de NADPH + H+ (toutes les deux provenant de la phase claire). Il y a donc consommation de 2 ATP et 2 NADPH + H+. Un sixième des trioses formés va entrer dans les réactions métaboliques de la plante. Ils seront principalement transformés en glucides (puis en saccharose pour le transport dans la sève élaborée, ou en amidon pour le stockage dans le chloroplaste). Il peut y avoir également formation de lipides (acides gras, glycérol) et d'acides aminés (alanine, glycine,...). Les cinq sixièmes des trioses restant dans le chloroplaste sont utilisés pour régénérer du RuDP. Toutes les réactions qui permettent cette régénération forment le cycle de Calvin. Au cours de ces réactions une molécule d'ATP est consommée, et certains produits intermédiaires peuvent être utilisés dans d'autres réactions métaboliques (glycolyse, voie des pentoses-phosphates, synthèse d'acides nucléiques).
Le bilan des réactions est donc la consommation de 3 molécules d'ATP et de 2 molécules de NADPH + H+ par molécule de CO2 incorporée soit 9 ATP et 6 NADPH + H+ consommés pour former un triose (phosphoglycéraldéhyde).
Les plantes en C3 présentent le phénomène de la photorespiration : l'oxygène entre en compétition avec la fixation du CO2. Dans le chloroplaste, la RUBISCO peut, en présence d'O2 transformer le RuDP en une molécule de PGA et une molécule d'acide phosphoglycolique : c’est la fonction oxygénase de la RUBISCO. Le PGA est incorporé dans le cycle de Calvin, tandis que l'acide phosphoglycolique, toxique pour la plante, sera transformé en glycine, au niveau d'un autre organite cellulaire : le peroxysome. Dans les mitochondries, deux molécules de glycine pourront se condenser pour donner une sérine, mais en entraînant la perte d'une molécule de CO2. Donc dans le cas de la photorespiration, il y a perte d'un CO2 au lieu d'avoir fixation d'un CO2. Cela entraîne une diminution du rendement de la photosynthèse. Ainsi la photosynthèse nette, qui est égale à la quantité de carbone fixé (photosynthèse brute) moins les pertes liées à la photorespiration et à la respiration n'est que de 20 à 25 mg de CO2 fixé par dm2 et par heure. Ce sont près de 6 mg de CO2 par dm2 et par heure qui sont perdus lors de la photorespiration, et 1 mg à cause de la respiration.
De plus, pour assimiler un gramme de CO2, de telles plantes perdent par transpiration plus de 600 grammes d'eau. C'est pourquoi certaines plantes, vivant dans des milieux plus difficiles doivent utiliser un système plus performant.
Ces plantes présentent une particularité anatomique. Au niveau des feuilles, on peut observer une organisation des tissus en couronne. Une première couronne de cellule, la gaine vasculaire, entourent les vaisseaux conducteurs. Une couronne plus externe est formée par les cellules du mésophylle. Les communications entre les cellules du mésophylle et de la gaine sont facilitées par la présence de nombreux plasmodesmes. L'épiderme est généralement recouvert d'une cuticule épaisse imperméable.
Dans les cellules du mésophylle, le CO2 s'associe au phospho-enol-pyruvate ou PEP pour former, grâce à l'action d'une enzyme spécifique la PEPcarboxylase, une molécule en C4, l'oxaloacétate. Selon les plantes, l'oxaloacétate va suivre des chaînes de réactions différentes mais qui aboutissent toutes au même résultat. Il y a formation d'une molécule de pyruvate dans les cellules de la gaine vasculaire et libération d'une molécule de CO2 dans les chloroplastes de ces cellules. Le CO2 suit alors le même trajet que chez les plantes C3 grâce à la présence de la RUBISCO dans ces chloroplastes. Le pyruvate permet de régénérer le PEP dans les cellules du mésophylle mais en consommant une molécule d'ATP.
Le bilan est ici une plus forte consommation d'ATP que chez les plantes C3 : 4 à 5 molécules d'ATP et 2 molécules de NADPH+H+ sont nécessaires pour incorporer une seule molécule de CO2. Mais ce n'est pas un facteur limitant pour la plante car l'ATP est présent en quantité suffisante.
Le rendement en matière sèche des plantes en C4 est beaucoup plus important que celui des plantes C3. D'une part, la PEPcarboxylase a une très forte affinité pour le CO2 (Km = 10-6 M)Celui-ci est présent dans la plante principalement sous forme dissoute (95%), or c'est sous cette forme que la PEPcarboxylase accepte le CO2, au contraire de la RUBISCO qui ne travaille qu'avec du CO2 sous forme gazeuse (avec un Km de 10-3 M seulement). La structure en couronne contribue à une forte augmentation de la concentration de CO2 dans les chloroplastes des cellules de la gaine vasculaire. La RUBISCO, en présence d’une forte teneur CO2, ne fixe pas l'oxygène, la photorespiration est donc fortement réduite. Enfin, le contact étroit entre la gaine vasculaire et les vaisseaux conducteurs rend l'exportation des produits formés plus efficace. La photosynthèse nette est ici de 40 à 60 mg de CO2 fixé par dm2 et par heure, près du double de la production des plantes en C3. La transpiration des plantes en C4 est limitée à 300 grammes d'eau pour un gramme de CO2 assimilé.
On ne retrouve pas de particularité anatomique chez ces plantes, bien qu'elles fonctionnent selon le même principe que les plantes en C4. Ici la différenciation est temporelle : les stomates sont fermés la journée pour protéger la plante de la dessiccation, le CO2 est fixé la nuit par la PEPcarboxylase. Tout se passe dans le parenchyme lacuneux où de l'acide malique s'accumule dans les vacuoles pendant la nuit conduisant une acidification notable du suc vacuolaire (acidification déjà remarquée par le genevois Nicolas-Théodore de Saussure au XIXème siècle) . Pendant la journée, cet acide malique sera dégradé en pyruvate et CO2. Le CO2 libéré dans les chloroplastes est pris en charge par la RUBISCO, selon les mêmes réactions que chez les plantes en C3 ou C4. La consommation d'ATP est encore plus importante que chez les C4 : 5 à 6 ATP et 2 molécules de NADPH+H+ sont nécessaires à l'incorporation d'une seule molécule de CO2. La production des plantes CAM est très variable. Le principale avantage de ce métabolisme est que le CO2 est « capté » la nuit pour former et stocker des molécules organiques. Le jour, ces molécules seront dégradées pour libérer du CO2. Ainsi les stomates peuvent rester fermés toute la journée pour limiter au maximum les pertes d’eau par transpiration.
Métabolisme des végétaux ; physiologie et biochimie – Gerhard Richter – PPUR, Lausanne, 1993 – ISBN 2-88074-231-5 – Voir en particulier le chapitre 3 intitulé « Autotrophie des organismes végétaux ».
Sur le web : http://www1.biologie.uni-hamburg.de/b-online/e00/default.htm
Houghton, R.A.,& G.M.Woodwell. Global climatic change. In Scientific American, 260(4), April 1989, pp.36-44.
On trouvera un guide de laboratoire en ligne à l’adresse : http://web.grinnell.edu/courses/bio/qubitmanual/ (ce guide est en anglais)
Groupes I.1 et II.1 : A
Groupes I.2 et II.2 : B
Vérifier les effets de la lumière et des variations de la concentration atmosphérique du gaz carbonique sur la fixation de celui-ci par les processus photosynthétiques
Ouvrir le fichier d’analyse « S151CO2 ».
Cliquer sur l’icône « LabPro » afin d’accéder à la configuration du boîtier de contrôle.
Vérifier que la connexion des différentes sondes est correctement effectuée.
Aller dans le menu « calibration » et cliquer sur le port correspondant à la sonde à CO2.
Effectuer le Zéro en montant la pompe à air, suivi de la colonne piégeant le CO2 puis de la colonne dessiccative.
Attendre la stabilisation et cliquer sur « Ok ». Régler ensuite la sonde pour une valeur connue : enlever la colonne piégeant le CO2 et fixer un sac d’air, contenant une quantité connue de CO2 (500 ppm) à l’entrée de la pompe à air. Attendre la stabilisation et cliquer sur « Ok ».
L’enregistrement est pré-réglé à 60 minutes. L’expérimentation pouvant durer plus longtemps, arrêter l’enregistrement avant 60 min, sauver les données et relancer un nouvel enregistrement.
Connecter la pompe à air à la chambre (vide mais fermée), en intercalant le régulateur de flux puis connecter la colonne dessicative à la sortie d’air de la chambre, et enfin, connecter la sonde à CO2. Régler le flux d’air à 500 ml/mn. Lancer l’enregistrement et attendre la stabilisation de la valeur de CO2. Ne pas souffler vers l’entrée d’air de la pompe car la sonde à CO2 est très sensible et cela pourrait fausser vos résultats. Pour la fenêtre enregistrant la consommation du CO2 régler l’axe des ordonnées (en cliquant sur l’axe avec la souris) 100 ppm au–dessous et 50 ppm au-dessus de la valeur observée. Ainsi pour une valeur observée de 400 ppm, régler la fenêtre des ordonnées entre 300 et 450 ppm
Sceller une feuille dans la chambre ; évaluer la superficie de la feuille grâce à la grille quadrillée (évaluer le nombre de carreaux couverts par la feuille et diviser la valeur par 4 pour avoir la superficie en cm2). Prendre garde de bien refermer hermétiquement la chambre d’échange gazeux.
Allumer la lumière à l’intensité maximale et suivre la consommation du CO2
Attendre que la valeur du CO2 soit stabilisée (une phase de déclin rapide suivie d’une phase plus lente, correspondant à la fermeture des stomates).
Eteindre ensuite la source lumineuse et patienter jusqu’à obtention d’une valeur stable. Suivre la variation des échanges gazeux en fonction de l’intensité lumineuse (20%, 40%, 60% et 80%). Attendre à chaque fois l’obtention d’une valeur stable.
Régler l’intensité lumineuse à ~80% et suivre les échanges gazeux en fonction de la concentration de CO2. Vous disposerez de plusieurs sacs contenant différentes concentrations de CO2. Monter successivement les sacs sur la pompe, en commençant par les concentrations les plus élevées.
Pour le dernier sac (CO2 = 0 ppm), suivre aussi les échanges gazeux à l’obscurité.
Arrêter l’enregistrement et sauver les données.
|
Conditions expérimentales |
Valeur en ppm (CO2) |
Valeur en μmol/m2/s |
Intensité lumineuse (μmol quanta/m2/s) |
|
Air labo seul (Réf) |
|||
|
Plante + Lumière |
|||
|
Plante + Obscurité |
|||
|
Intensité 20% |
|||
|
Intensité 40% |
|||
|
Intensité 60% |
|||
|
Intensité 80% |
|||
|
CO2 1000 ppm |
|||
|
CO2 500 ppm |
|||
|
CO2 360 ppm (air) |
|||
|
CO2 200 ppm |
|||
|
CO2 0 ppm |
|||
|
CO2 0 ppm + Obs |
Ouvrir le ou les fichiers de données.
Convertir les valeurs en μmol CO2/m2/s pour remplir le tableau :
Déterminer la consommation de CO2 (dCO2) en ppm (référence-observé)
Appliquer la formule suivante : dCO2/{22,413 [(273+T)/273]} pour obtenir la valeur de CO2 consommé en μmol/l (T étant la température)
Multiplier la valeur obtenue par le flux en l/s pour obtenir la quantité de CO2 consommé en μmol/s.
Convertir la surface de la feuille en m2 et exprimer ensuite la consommation du CO2 par unité de surface (μmoles CO2/m2/s)
Vérifier les effets de la lumière (intensité, qualité) sur la production d’oxygène par la photosynthèse
Ouvrir le fichier d’analyse « Ph1 ».
Cliquer sur l’icône « LabPro » afin d’accéder à la configuration du boîtier de contrôle.
Vérifier que la connexion des différentes sondes est correctement effectuée.
Lancer l’enregistrement. Placer une feuille dans la chambre des échanges gazeux et bien la refermer hermétiquement. Evaluer la superficie de la feuille grâce à la grille quadrillée (évaluer le nombre de carreaux couverts par la feuille et diviser la valeur par 4 pour avoir la superficie en cm2).
Ouvrir une des entrées d’air de la chambre et souffler dedans grâce à une paille pendant quelques secondes. Ainsi la chambre contiendra un air enrichi en CO2.
Placer un Becher rempli d’eau au-dessus de la chambre, dans le cas où la source de lumière est une lampe halogène (évite l’échauffement dans la chambre).
Allumer la lumière à l’intensité maximale et suivre la production de O2.
Attendre l’obtention d’une pente bien droite (une phase de déclin rapide suivie d’une phase plus lente, correspondant à l’ouverture des stomates).
Eteindre ensuite la source lumineuse et patienter jusqu’à obtention d’une pente stable. Suivre la variation des échanges gazeux en fonction de l’intensité lumineuse (20%, 40%, 60% et 80%). Attendre à chaque fois la stabilisation.
Tout en conservant l’intensité lumineuse à 80%, placer l’acétate de couleur bleue entre la source lumineuse et la feuille. Attendre de nouveau la stabilisation. Effectuer la même opération avec les autres acétates de couleur.
Pour le dernier sac (CO2 = 0 ppm), suivre les échanges gazeux à l’obscurité.
Arrêter l’enregistrement et sauver les données.
|
Conditions expérimentales |
Valeur CO2 ppm/mn ou O2 en %/mn |
Valeur en μmol/m2/s |
Intensité lumineuse (μmol quanta/m2/s) |
|
Air labo seul (Réf) |
|||
|
Air soufflé (exp) |
|||
|
Plante + Obscurité |
|||
|
Plante + Lumière |
|||
|
Intensité 20% |
|||
|
Intensité 40% |
|||
|
Intensité 60% |
|||
|
Intensité 80% |
|||
|
Acétate Rouge |
|||
|
Acétate Bleu |
|||
|
Acétate Vert |
Il est important de noter la concentration initiale de O2 de l’air (~20,70%). Quand vous soufflez de l’air dans la chambre d’échange gazeux, l’air est enrichi en CO2 aux dépends de l’O2. En considérant que la concentration de CO2 normale de l’air est d’environ 0.05 %, si la concentration de O2 de l’air soufflé est de 18 %, la concentration de CO2 est déterminée par l’opération suivante :
(20,70 – 18) + 0,05 = 2,75 %
Quand au cours de l’expérimentation, la concentration de O2 atteint 20,15%, en 10 minutes la concentration de CO2 est déterminée par la même opération :
(20,70 – 20,15) + 0,05 = 0,65 %
Cette valeur équivaut à 6500 ppm de CO2 produits en 10 minutes, soit 650 ppm/mn. Reporter cette valeur dans le tableau.
Déterminer la consommation de CO2 en ppm/mn.
Appliquer la formule suivante : CO2/{22,413 [(273+T)/273]} pour obtenir la valeur de CO2 consommé en μmoles/l/mn (T étant la température en °C)
Sachant que le volume fixe de la chambre est de 0,047 litres, convertir cette valeur en μmoles/s
Convertir la surface de la feuille en m2 et exprimer ensuite la consommation du CO2 par unité de surface (μmoles CO2/m2/s)
Tracer la courbe de consommation de CO2 en fonction de la concentration.
Discuter des résultats obtenus avec les plantes en C3 et en C4
Quel est l’effet physiologique de l’augmentation de l’intensité lumineuse ?
Que se produit-il lorsqu’on filtre la lumière avec des acétates colorés ?
L'homme et les animaux ont besoin d'azote sous forme organique pour leur alimentation, tandis que les végétaux sont capables d'assimiler l'azote minéral sous forme de nitrates (NO3-) et d'ammoniaque (NH3).
L'azote atmosphérique est la seule réserve d'azote accessible de la biosphère. Pratiquement illimitée, cette réserve (4.1015 tonnes) n'est pas directement utilisable par les végétaux et les animaux. On estime que l’azote organique de notre planète (4.1013 tonnes) dérive de l’atmosphère grâce à la fixation biologique de l’azote par les bactéries.
La découverte de la fixation l'azote par les Légumineuses remonte au 19ème siècle. Il a fallu plus de cinquante ans, entre les travaux du français Jean-Baptiste Boussingault et ceux des allemands Hermann Hellriegel et Hermann Wilfarth, pour établir, en 1886, que l'aptitude des légumineuses à utiliser l'azote de l'air était due à la présence de nodosités, sur les racines, induites par des "ferments" contenus dans le sol. Cette découverte fut rapidement suivie par l'isolement des Rhizobium par Beijerinck, puis de plusieurs autres bactéries fixatrices d'azote, non associées aux plantes, comme Azotobacter et Clostridium pasteurianum.
La fixation biologique de l'azote est estimée actuellement à près de 200 millions de tonnes par an, soit plus de deux fois la production d'engrais azotés par synthèse industrielle. A l'état naturel, les fixateurs d'azote peuvent être associés à des plantes (symbioses vraies ou associatives) ou demeurer à l'état libre dans le sol et les eaux. Les symbioses vraies (Rhizobium-légumineuses par exemple) sont les plus efficaces. Les légumineuses à elles seules sont responsables de près de 30 % de la fixation d’azote annuelle (avec des taux annuels de 30 à 300 kg N fixés par hectare).
On connaît plus de 20.000 espèces de légumineuses et sur 3.000 espèces examinées pour la propriété à former des nodosités, 90 % portaient des nodosités sur les racines contenant des bactéries appartenant au genre Rhizobium ou Bradyrhizobium. Récemment certaines souches, appartenant au genre Azorhizobium ont été isolées à partir de nodules localisés sur la tige de la plante hôte, comme Sesbania rostrata. Cette bactérie est également capable d'induire la formation de nodosités sur les racines.
La symbiose Rhizobium-Légumineuses est le résultat d'une interaction hautement spécifique entre le partenaire bactérien et le partenaire végétal. La morphogenèse de la nodosité est induite par un échange de signaux entre la plante et la bactérie. Dans la nodosité la bactérie est sous une forme non proliférante, le bactéroïde, qui est la forme capable de fixer l'azote. L'azote fixé, sous forme d'ammoniaque, est sécrété vers la plante qui en échange fournit les substrats carbonés et un système de transport d’oxygène (léghémoglobine) nécessaires au bon développement des bactéries.
Le complexe enzymatique: Les bactéries fixatrices d'azote convertissent l'azote moléculaire en ammoniaque grâce à un complexe enzymatique, appelé nitrogénase, formé de deux métalloprotéines qui comportent trois groupements prosthétiques différents. Les deux métalloprotéines composant la nitrogénase sont le plus souvent appelées Protéine I et Protéine II. C'est la Protéine I qui contient le cofacteur à fer et à molybdène appelé FeMo-co au niveau duquel se produit la réduction du substrat. Toutefois, l'atome de molybdène n'est pas le site direct de la réduction de l'azote comme on l'a longtemps pensé.
En présence d'azote moléculaire comme substrat, la réaction catalysée par la nitrogénase conduit toujours, en plus de l'ammoniaque, à la production d'hydrogène par réduction de protons. Le complexe catalyse également la réduction d'analogues stériques de l'azote, comme l'acétylène qui est réduit en éthylène. L'éthylène produit est facilement dosable par chromatographie en phase gazeuse. Cette propriété est souvent utilisée en laboratoire pour doser l’activité nitrogénase, en utilisant un mélange gazeux à 10% d’acétylène, mais assez délicat en raison du caractère explosif de l’acétylène. Un meilleur moyen consiste à doser l’hydrogène produit lors de la réaction.
Ce test n’est pas toujours valable car plusieurs espèces de Rhizobium et Bradyrhizobium possèdent une hydrogénase capable de ré-oxyder l’hydrogène produit lors de la réaction de la nitrogénase, mais ce n’est pas la majorité. Les expériences suivantes sont réalisées avec des plantes en symbiose avec une souche de Bradyrhizobium qui ne possède pas cet enzyme : dans ce cas, l’hydrogène produit diffuse librement dans le sol.

En présence de N2, trois paires d’électrons sont utilisées pour réduire le N2 en 2 NH3 alors qu’une paire d’électrons est utilisée pour réduire deux protons en H2. En absence de N2 (on utilise un gaz inerte, l’Hélium), les huit électrons seront uniquement utilisés pour la réduction des protons, donc produire 4 molécules de H2. Le dosage du H2 en présence de N2 donne l’activité nitrogénase apparente (ANA), tandis qu’en absence de N2, on observe l’activité nitrogénase totale (ANT). On obtient ainsi les équations suivantes :
Taux de fixation de N2 = (ANT - ANA) / 3
Coefficient d’allocation des électrons (CAE) = 1 – (ANA/ANT)
Métabolisme des végétaux ; physiologie et biochimie – Gerhard Richter – PPUR, Lausanne, 1993 – ISBN 2-88074-231-5 – Voir en particulier le chapitre 9 intitulé « Composés azotés : acides aminés et nucléotides ».
Sur le web : http://www1.biologie.uni-hamburg.de/b-online/e00/default.htm
On trouvera un guide de laboratoire en ligne à l’adresse : http://web.grinnell.edu/courses/bio/qubitmanual/ (ce guide est en anglais)
Groupe I.1 : A & B ; Groupe I.2 :A & C
Groupe I.2 : A & D ; Groupe II.2 : A & E
Vérifier la stœchiométrie entre le nombre d’électrons transférés et le nombre de protons (H2) produits

A- Colonne de dessicant
B-http://web.grinnell.edu/courses/bio/qubitmanual/Information/nitrogen/O2SENSOR.HTM Amplificateur du senseur à O2
C- Senseur à oxygène
E- Pompe à air
G- Régulateur de flux
J- Plante en pot
Un sac de mélange He:O2 (80:20)
Vérifier que le matériel est monté correctement comme indiqué sur le schéma.
Fixer la colonne de dessicant directement sur la sortie de la pompe à air.
Lancer le programme Logger Pro et charger le fichier « N2FixO2 » (dossier « Viernier Software »)
Deux graphiques apparaissent sur l’écran, celui du haut présente l’enregistrement des données du senseur H2 et celui du bas, le senseur à O2.
Ouvrir uniquement la fenêtre du senseur H2; en abscisse est indiqué le temps, calibré à 60 min pour cette expérience. Si l’expérience dépasse 60 min, les données ne seront pas enregistrées au-delà, donc il faut arrêter l’enregistrement, sauver les données et relancer un nouvel enregistrement.
Fixer à l’entrée de la pompe le sac contenant le mélange He:O2 (80:20) et régler le flux d’air à 200 ml/min. Lancer l’enregistrement.
Attendre environ 5 min que la valeur se stabilise et noter ce qui sera le Zéro pour cette expérience.
Enlever le sac d’argon et laisser l’enregistrement courir quelques minutes pour obtenir le zéro, pour les expériences réalisées avec l’air.
Intercaler la plante entre la pompe à air et la colonne de dessicant et augmenter le flux d’air à 500 ml/min (s’assurer de l’étanchéité du pot au niveau du collet de la plante).
Observer l’augmentation de la production et attendre que cette valeur soit stabilisée.
Réduire le flux d’air à 200 ml/min et observer la variation de l’enregistrement des données du senseur H2. attendre de nouveau que la valeur soit stabilisée.
Augmenter de nouveau le flux à 500 ml/min et attendre la stabilisation.
Attacher à l’entrée de la pompe le sac contenant le mélange He:O2 (80:20) et observer les modifications de l’enregistrement. Attendre l’obtention d’une valeur stable.
Arrêter l’enregistrement et sauver les données obtenues.
Ouvrir le fichier de données.
Sélectionner « Examine » dans le sous-menu « analyze ». Ainsi, vous pouvez sélectionner des régions de la courbe d’enregistrement et les valeurs expérimentales s’affichent dans une fenêtre.
Les valeurs sont données en volts et doivent être converties comme suit :
H2 production (μmol/h) = Voltage x K x Flux
Attention à bien convertir le Flux en litre par heure. La valeur K est différente suivant les gaz utilisés :
K = 0,75 μmol/l par volt dans l’air
K = 1,0 μmol/l par volt dans He:O2
Ne pas oublier de soustraire la valeur du Zéro correspondante pour chaque calcul.
H2 Zéro avec l’air = …. Volts
H2 Zéro avec He:O2 = …. Volts
|
Flux (L/heure) |
H2 dans air (Volts) |
H2 dans He:O2 (Volts) |
H2 Zéro (Volts) |
K |
H2 produit (μmol/h) |
Calculer les valeurs de l’activité nitrogénase apparente (ANA) et totale (ANT) et utiliser ces valeurs pour obtenir le coefficient d’allocation d’électrons (CAE) de la plante. Pourquoi cette valeur ne peut-elle pas excéder 0.75 ?
Que se passe-t-il quand on modifie le flux d’air, pourquoi le voltage est-il affecté ? Cela modifie-t-il l’ANA ?
Sachant que 2 électrons sont nécessaires pour réduire 2 protons en H2, et que 6 électrons sont nécessaires pour réduire une molécule de N2 en 2 NH3, calculer en utilisant les valeurs observées le taux de fixation de N2 dans la plante en μmol fixé par heure.
Les légumineuses ont un CAE maximal de 0,75. Quel serait l’effet sur la plante si cette valeur était fortement diminuée ?
Utiliser la méthode mesure du H2 produit pour étudier les effets de la température sur la capacité de fixation de l’azote par les nodules
Deux plantes en pot
Une sonde de température
Un bac de glace
Un bain-marie à 50°C
Un bain-marie à 70°C
Enlever l’électrode à O2, insérer la sonde à température dans le connecteur « CH2 » sur le boîtier relié à l’ordinateur.
Fixer la pompe à air directement sur la colonne de dessicant et ajuster le flux à 200 ml/min.
Ouvrir le fichier « N2FixTemp ».
Lancer l’enregistrement et attendre quelques minutes pour obtenir le Zéro.
Plonger la sonde de température au cœur d’un pot. Prendre soin de bien faire l’étanchéité autour de la sonde et du collet de la plante. On peut vérifier l’étanchéité du système en branchant la pompe à air sur l’entrée du pot et en plongeant la sortie d’air dans un récipient rempli d’eau.
Monter le pot entre la pompe à air et la colonne de dessicant.
Observer les variations du voltage et attendre la stabilisation (10-15 min).
Une fois le voltage stabilisé, plonger le pot dans le bac de glace. Suivre la diminution du voltage en parallèle à la diminution de la température des racines, jusqu’à atteindre une température de 10°C.
Changer la plante sur le montage, attendre la stabilisation du voltage et placer le pot dans le bain-marie à 50°C.
Suivre l’évolution du voltage en fonction de l’augmentation de la température.
Quand la température est dans les environs de 40°C, placer alors le pot dans le bain-marie à 70°C.
Continuer l’observation jusqu’à ce que la température enregistrée soit au-delà de 50°C et que le voltage diminue sensiblement. Arrêter l’enregistrement et sauver les données.
Ouvrir le fichier de données.
Sélectionner « Examine » dans le sous-menu « analyze ». Ainsi, vous pouvez sélectionner des régions de la courbe d’enregistrement et les valeurs expérimentales s’affichent dans une fenêtre.
Les valeurs sont données en volts et doivent être converties comme suit :
H2 production (μmol/h) = Voltage x K x Flux
K = 0,75 μmol/l par volt dans l’air
Ne pas oublier de soustraire la valeur du Zéro correspondante pour chaque calcul.
Flux = litre/heure
H2 Zéro = Volts
|
Température (°C) |
Production H2 (Volts) |
H2 Zéro (Volts) |
Activité nitrogénase (μmol H2/heure) |
|
Température (°C) |
Production H2 (Volts) |
H2 Zéro (Volts) |
Activité nitrogénase (μmol H2/heure) |
Le Q10 est le rapport de l’activité à une température donnée sur l’activité à une température 10°C inférieure. Ainsi une réaction qui est deux fois plus active avec une augmentation de température de 10°C aura un Q10 = 2. Rapporter sur un graphique les valeurs observées (activité en μmol H2/heure par rapport à la température). Quelle est la température optimale de la réaction ? Quel est son Q10 ? Le Q10 est-il différent s’il est calculé dans les basses températures en comparaison avec les hautes températures ?
Pourquoi deux plantes différentes ont-elles été utilisées lors de cette expérience ? Qu’aurait-on pu observer si la même plante avait été exposée à de basses températures puis soumise à de fortes températures ?
Utiliser la méthode mesure du H2 produit pour étudier les effets de la tension d’oxygène sur la fixation de l’azote par les nodules sachant que l’oxygène affecte fortement la nitrogénase.
Deux plantes en pot
Sonde à O2
Deux sacs contenant un mélange N2:O2 (70:30)
Une cuvette pour placer les nodule
Brancher la pompe à air directement sur la colonne et régler le flux à 200 ml/min.
Charger le fichier « N2FixO2 » et lancer l’enregistrement. L’enregistrement de l’O2 doit indiquer 20,9%, si ce n’est pas le cas, modifier la sensibilité de l’amplificateur pour obtenir cette valeur. Attendre quelques minutes que la valeur pour H2 soit stabilisée.
L’expérience est en deux parties : une partie sur des racines intactes et l’autre partie sur des nodules isolés.
Attacher le sac du mélange N2:O2 (70:30) sur la pompe et mesurer à nouveau le Zéro.
Détacher et sceller le sac de gaz. Intercaler la plante en pot entre la pompe et la colonne (vérifier l’étanchéité du pot) et augmenter le flux d’air à 500 ml/min. Attendre la stabilisation du voltage (15-20 min) et noter la valeur.
Attacher le sac contenant le mélange N2:O2 (70:30) à l’entrée de la pompe. Attendre la stabilisation du voltage (20-30 min) et arrêter l’enregistrement. Sauver les données.
Remplacer la plante utilisée lors de la manipulation précédente par une deuxième à disposition. Relancer l’enregistrement de la production de H2 et attendre que la valeur se stabilise (15-20 min).
Enlever la plante du montage et rapidement, sortir la plante du pot et récupérer tous les nodules présents sur les racines. Les mettre dans la cuvette avec du papier imbibé d’eau.
Monter la cuvette entre la pompe à air et la colonne de dessicant et régler le flux d’air à 200 ml/min. Attendre que la valeur se stabilise (10-15 min).
Attacher le sac du mélange N2:O2 (70:30) à l’entrée de la pompe et observer l’évolution de la production de H2, jusqu’à obtention d’une valeur stable (10-15 min).
Arrêter l’enregistrement et sauvegarder les donner.
Ouvrir le fichier de données. Sélectionner « Examine » dans le sous-menu « analyze ». Ainsi, vous pouvez sélectionner des régions de la courbe d’enregistrement et les valeurs expérimentales s’affichent dans une fenêtre.
Les valeurs sont données en volts et doivent être converties comme suit :
H2 production (μmol/h) = Voltage x K x Flux
Soustraire la valeur du Zéro correspondante pour chaque calcul. Attention à bien convertir le Flux en litre par heure. La valeur K est différente suivant les gaz utilisés :
K = 0,75 μmol/l par volt dans l’air
K = 0,95 μmol/l par volt dans N2:O2 (70:30)
H2 Zéro avec l’air = ….. Volts
H2 Zéro N2:O2 (70:30) = ….. Volts
Valeurs observées (en Volts)
|
Activité stable à 20%O2 (Volts) |
Activité min. à 30%O2 (Volts) |
Activité max. à 30%O2 (Volts) |
|
|
Plante 1 |
|||
|
Plante 2 |
Non effectué |
Non effectué |
|
|
Nodules isolés |
En utilisant les valeurs observées, calculer la production de H2 et reporter ces valeurs dans le tableau ci-dessous. Ne pas oublier de soustraire les « Zéros » appropriés, d’utiliser les valeurs de K adéquates ainsi que le flux d’air correspondant aux conditions expérimentales.
Valeurs calculées (en μmolH2/heure)
|
Activité stable à 20%O2 (μmolH2/heure) |
Activité min. à 30%O2 (μmolH2/heure) |
Activité max. à 30%O2 (μmolH2/heure) |
|
|
Plante 1 |
|||
|
Plante 2 |
Non effectué |
Non effectué |
|
|
Nodules isolés |
Comment évolue l’activité nitrogénase quand on augmente la concentration en O2 ?
L’inhibition de la nitrogénase est-elle irréversible ? Commenter et établir une hypothèse pour expliquer les résultats observés.
Comment les racines produisent-elles l’énergie dont elles ont besoin ? Quelle en est la conséquence quand on isole les nodules des racines ? Expliquer ce qu’on observe sur des nodules isolés quand on augmente la concentration en O2.
Les deux prochaines expériences vous permettent d’examiner deux corrélations physiologiques qui affectent la fixation de l’azote ; tout d’abord en examinant l’effet de l’apport d’un engrais azoté (nitrate)
Une plante contrôle, cultivée dans un milieu pauvre en nitrates
Une plante cultivée dans un milieu riche en nitrates
Sonde à O2
Deux sacs contenant un mélange N2:O2 (70:30)
Une cuvette pour placer les nodules
Brancher la pompe à air directement sur la colonne et régler le flux à 200 ml/min.
Charger le fichier « N2FixO2 » et lancer l’enregistrement. L’enregistrement de l’O2 doit indiquer 20,9%, si ce n’est pas le cas, modifier la sensibilité de l’amplificateur pour obtenir cette valeur. Attendre quelques minutes que la valeur pour H2 soit stabilisée.
L’expérience est en deux parties : une partie sur des racines intactes et l’autre partie sur des nodules isolés.
Attacher le sac du mélange N2:O2 (70:30) sur la pompe et mesurer à nouveau le Zéro.
Détacher et sceller le sac de gaz. Intercaler la plante contrôle entre la pompe et la colonne (vérifier l’étanchéité du pot) et augmenter le flux d’air à 500 ml/min. Attendre la stabilisation du voltage (15-20 min) et noter la valeur.
Attacher le sac contenant le mélange N2:O2 (70:30) à l’entrée de la pompe. Attendre la stabilisation du voltage (20-30 min) et arrêter l’enregistrement. Sauver les données.
Enlever la plante contrôle du montage et répéter les mêmes opérations avec la plante cultivée dans un milieu riche en nitrates (enregistrement de la production de H2 en présence d’air, puis en présence du mélange à 30% de O2).
Arrêter l’enregistrement et sauvegarder les donner.
Isoler séparément tous les nodules des deux plantes et les peser.
Ouvrir le fichier de données. Sélectionner « Examine » dans le sous-menu « analyze ». Ainsi, vous pouvez sélectionner des régions de la courbe d’enregistrement et les valeurs expérimentales s’affichent dans une fenêtre.
Les valeurs sont données en volts et doivent être converties comme suit :
H2 production (μmol/h) = Voltage x K x Flux
Soustraire la valeur du Zéro correspondante pour chaque calcul. Attention à bien convertir le Flux en litre par heure. La valeur K est différente suivant les gaz utilisés :
K = 0,75 μmol/l par volt dans l’air
K = 0,95 μmol/l par volt dans N2:O2 (70:30)
|
H2 Zéro avec l’air = ….. Volts |
Nodules contrôles = …. g |
|
H2 Zéro N2:O2 (70:30) = ..… Volts |
Nodules traités = …. g |
Valeurs observées (en Volts)
|
Activité stable à 20%O2 (Volts) |
Activité min. à 30%O2 (Volts) |
Activité max. à 30%O2 (Volts) |
|
|
Plante contrôle |
|||
|
Plante traitée |
En utilisant les valeurs observées, calculer la production de H2 et reporter ces valeurs dans le tableau ci-dessous. Ne pas oublier de soustraire les « Zéros » appropriés, d’utiliser les valeurs de K adéquates ainsi que le flux d’air correspondant aux conditions expérimentales.
Valeurs calculées (en μmolH2/heure)
|
Activité stable à 20%O2 (μmolH2/heure) |
Activité min. à 30%O2 (μmolH2/heure) |
Activité max. à 30%O2 (μmolH2/heure) |
|
|
Plante contrôle |
|||
|
Plante traitée |
Comment évolue l’activité nitrogénase quand on augmente la concentration en O2 ?
L’inhibition de la nitrogénase est-elle irréversible ? Commenter et établir une hypothèse pour expliquer les résultats observés.
Que peut-on observer pour les plantes traitées aux nitrates ?
Comparer les activités spécifiques (μmol H2/g nodules/heure) de la nitrogénase des plantes contrôles et des plantes cultivées dans un milieu riche en nitrates.
Comparer les résultats des plantes traitées aux nitrates avec ceux obtenus avec les nodules isolés (poste voisin) : quel est le constat ? Comparer vos hypothèses.
La photosynthèse fournit la plante et les nodules en éléments nutritifs; vous allez moduler ce facteur en agissant sur la lumière et la quantité de feuilles
Une plante dont on a coupé la tige trois heures avant l’expérience
Une plante ayant subi 24h d’obscurité.
Sonde à O2
Deux sacs contenant un mélange N2:O2 (70:30)
Une cuvette pour placer les nodules
Brancher la pompe à air directement sur la colonne et régler le flux à 200 ml/min.
Charger le fichier « N2FixO2 » et lancer l’enregistrement. L’enregistrement de l’O2 doit indiquer 20,9%, si ce n’est pas le cas, modifier la sensibilité de l’amplificateur pour obtenir cette valeur. Attendre quelques minutes que la valeur pour H2 soit stabilisée.
L’expérience est en deux parties : une partie sur des racines intactes et l’autre partie sur des nodules isolés.
Attacher le sac du mélange N2:O2 (70:30) sur la pompe et mesurer à nouveau le Zéro.
Détacher et sceller le sac de gaz. Intercaler la plante ayant subi 24h d’obscurité entre la pompe et la colonne (vérifier l’étanchéité du pot) et augmenter le flux d’air à 500 ml/min. Attendre la stabilisation du voltage (15-20 min) et noter la valeur.
Attacher le sac contenant le mélange N2:O2 (70:30) à l’entrée de la pompe. Attendre la stabilisation du voltage (20-30 min) et arrêter l’enregistrement. Sauver les données.
Enlever la plante du montage et répéter les mêmes opérations avec la plante décapitée (enregistrement de la production de H2 en présence d’air, puis en présence du mélange à 30% de O2).
Arrêter l’enregistrement et sauvegarder les donner.
Isoler séparément tous les nodules des deux plantes et les peser.
Récupérer les valeurs obtenues pour une plante contrôle auprès de vos voisins.
Ouvrir le fichier de données. Sélectionner « Examine » dans le sous-menu « analyze ». Ainsi, vous pouvez sélectionner des régions de la courbe d’enregistrement et les valeurs expérimentales s’affichent dans une fenêtre.
Les valeurs sont données en volts et doivent être converties comme suit :
H2 production (μmol/h) = Voltage x K x Flux
Soustraire la valeur du Zéro correspondante pour chaque calcul. Attention à bien convertir le Flux en litre par heure. La valeur K est différente suivant les gaz utilisés :
K = 0,75 μmol/l par volt dans l’air
K = 0,95 μmol/l par volt dans N2:O2 (70:30)
|
Nodules contrôles = …. g |
|
|
H2 Zéro avec l’air = ….. Volts |
Nodules 24h obs = ….. g |
|
H2 Zéro N2:O2 (70:30) = ..... Volts |
Nodules décap. = ….. g |
Valeurs observées (en Volts)
|
Activité stable à 20%O2 (Volts) |
Activité min. à 30%O2 (Volts) |
Activité max à 30%O2 (Volts) |
|
|
Plante contrôle |
|||
|
Plante 24h obscurité |
|||
|
Plante décapitée |
En utilisant les valeurs observées, calculer la production de H2 et reporter ces valeurs dans le tableau ci-dessous. Ne pas oublier de soustraire les « Zéros » appropriés, d’utiliser les valeurs de K adéquates ainsi que le flux d’air correspondant aux conditions expérimentales.
Valeurs calculées (en μmolH2/heure)
|
Activité stable à 20%O2 (μmolH2/heure) |
Activité min. à 30%O2 (μmolH2/heure) |
Activité max à 30%O2 (μmolH2/heure) |
|
|
Plante contrôle |
|||
|
Plante 24h obscurité |
|||
|
Plante décapitée |
Comment évolue l’activité nitrogénase quand on augmente la concentration en O2 ?
L’inhibition de la nitrogénase est-elle irréversible ? Commenter et établir une hypothèse pour expliquer les résultats observés.
Comparer les activités spécifiques (μmol H2/g nodules/heure) de la nitrogénase des différentes plantes utilisées avec des plantes contrôles.
Comparer les résultats obtenus suite aux différents traitements (obscurité, décapitation et augmentation de la teneur en O2) avec ceux deux postes voisins. Commenter.
A première vue, les expériences groupées dans cette manipulation et le matériel biologique employé peuvent paraître disparates. En fait, il ne sera question ici que de certains types d'échanges cellulaires (osmose : eau ; perméation : ions) établis entre le vivant et son milieu. Quant aux sujets d'analyse (tubercules, épidermes, protoplastes..), ils ont été choisis parce qu'ils permettaient des expériences simples et de courte durée.
Quelques notions essentielles - largement développées au cours - seront brièvement et d'une façon élémentaire résumées ici. Cette introduction sommaire rendra accessible cette manipulation même à ceux qui n'auraient pas encore suivi les chapitres concernés.
Le potentiel osmotique, désignée par le symbole Ψo, est fonction de la température, de la concentration de la substance dissoute, et de ses caractéristiques de dissociation.
Ψo = R . T . c . i
Ψo : potentiel osmotique.. Il s'agit d'une forme du potentiel hydrique exprimé en atmosphères, en bars ou en pascals. (1 atmosphère = 1,013 bars. 1 bar = 106 pascals)
R : constante des gaz parfaits (8.316 . 107 ergs / mol .°K). Elle correspond au produit de la constante de Boltzmann par le Nombre d'Avogadro.
= 22.4 / 273 atm / M.°K)
T : température absolue (donnée en °K).
c : concentration molaire du corps dissous, exprimée en osmole.
i : coefficient d'ionisation (appelé parfois coefficient isotonique)
i = 1 + (γ - 1) . α
γ : nombre de ions présents dans une molécule du composé dissous
α : degré d'ionisation du composé dissous
Pour un non-électrolyte, i=1
Pour un électrolyte, calculons, à titre d'exemple, la valeur de π d'une solution
M / 3 de KCl dissociée aux 3 / 4 à 20 °C :
KCl K+ + Cl-
γ= 2 et α = 0.75 donc i = 7 / 4
avec RT (à 20°C) = 2.437*10-3 m3.MPa .mol-1
Ψo = 2.437*10-3 m3.MPa .mol-1 * 103 / 3 *7 / 4 MPa = 1.4 MPa
La notion de potentiel hydrique a été largement discutée au cours. On en trouvera les bases aux pages 24 à 31 de l'ouvrage de G. Richter "Métabolisme des végétaux".
Pour éclaircir des problèmes de terminologie, nous donnerons le tableau suivant :
|
Milieu intérieur |
Milieu extérieur |
ΔΨ = ΨINT - ΨEXT |
||
|
Iso-osmotique |
Isotonique |
équilibre |
ΔΨ = 0 |
|
|
Hypo-osmotique |
Hypertonique |
plasmolyse * |
ΔΨ > 0 |
* l'eau sort |
|
Hyper-osmotique |
Hypotonique |
turgescence ** |
ΔΨ < 0 |
** l'eau entre |
Rappelons aussi que l'anatonose permet à la cellule de diminuer son potentiel hydrique en dépolymérisant ses réserves hydrosolubles. L'épictèse est aussi une forme d'adaptation osmotique qui se traduit également par une diminution de son potentiel hydrique, mais à la suite d'une perméation d'ions exogènes.
La perméation est le phénomène de passage (ex. la perméation active du K+), alors que la perméabilité est la propriété de laisser passer (ex. le plasmalemme est perméable au Ca++). Si l'on évalue la quantité (q) d'un ion, par exemple, qui est entré dans un organe (disons une racine), on constate que l'absorption n'est que partiellement bloquée si le métabolisme est réduit. La première expérience nous donne la perméation globale (PG) du ion, la seconde sa perméation passive (PP), d'où l'on peut déduire sa perméation active (PA) :
PA = PG - PP et PG = PA + PP
Ce graphique fait apparaître une quantité particulière (e) qui change avec la concentration (c) du ion dans le milieu :
![]() donc e = K . c
donc e = K . c
On notera que la dimension de K est celle d'un volume ... c'est celui de l'espace libre, caractérisé - dans la cellule - par le passage à la fois actif et passif du ion considéré.
La perméation passive dépend du potentiel thermodynamique (entraînement : potentiel hydrique; diffusion: potentiel électrochimique), de l'électro-osmose et des équilibres de Gibbs et Donnan. La perméation active implique la participation de transporteurs (cytochromes..) et d'enzymes (perméases). Il faut encore mentionner la perméation sélective qui permet à la cellule de "choisir" les ions par une sorte de "discrimination" dont les mécanismes sont loin d'être tous élucidés.
(peut être modifié selon les indications de l'assistant)
Groupe I.1: A et Ca Groupe II.1 : A et Cb
Groupe I.2 : D Groupe II.2 : E
On déterminera le pouvoir osmotique relatif d'un tubercule de pomme de terre en réalisant le montage suivant :

On procédera de la façon suivante :
- Ménager un trou T dans le tubercule. A cet effet :
- Enfoncer (4-6 cm) un emporte-pièce dans le sens longitudinal du tubercule.
- Pour faire sortir le cylindre de pomme de terre: enfoncer (3-5 cm) un deuxième emporte-pièce de diamètre juste inférieur; faire subir aux deux emporte-pièces un mouvement simultané de rotation.
- Rincer le trou à l'eau et enlever les déchets.
- Estimer le volume V et la surface S du cylindre T.
- Remplir complètement T avec une solution Sl de saccharose 0,6 M, contenant une substance colorée (éosine) .
- Introduire le bouchon B, muni d'un capillaire TC. Au cours de cette opération le liquide Sl doit monter dans TC. Vérifier que toutes les bulles d'air sortent.
- Bien fixer le bouchon B et laisser le liquide Sl déborder par l'extrémité de TC si nécessaire.
- Diminuer le niveau N du liquide dans TC, en relevant le bouchon B.
- Fixer le tout sur un support et mesurer la montée du liquide dans le tube TC, toutes les 30 sec, pendant les 5 premières min, puis toutes les min pendant 5 min, et enfin après 20 min.
- En utilisant un autre tubercule, répéter la même opération avec une solution S2 (saccharose 0,2 M).
- Ramener les résultats à la surface S des cylindres T.
Reporter les valeurs ainsi obtenues sur un graphique.
- Interpréter et discuter les courbes obtenues.
Ce type d'osmomètre détermine le point de congélation des solutions aqueuses. L'abaissement du point de congélation par rapport à l'eau distillés donne une mesure directe de la concentration osmolaire. L'eau pure se solidifie à 0°C; une solution aqueuse avec une osmolarité de 1 osmole par kilo d'eau (Osml / kg) se solidifie à -1.858°C.
On procédera d'abord au calibrage de l'équipement, puis l'on effectuera les mesures des solutions à analyser dans les différentes expériences (voir C et suivant)
1. Mettre l’appareil sous tension à l’aide de l’interrupteur. Attendre environ 5 min.
2. Pipeter 100 μl d’eau distillée dans un tube Eppendorf.
3. Fixer le tube sur la tête de mesure.
4. Abaisser la tête de mesure pour introduire le tube Eppendorf dans le compartiment réfrigéré.
5. La température de surfusion est atteinte après 2 min environ.
6. Lorsque l’affichage atteint la valeur de -1000, l’aiguille catalyseuse se déplace automatiquement et plonge dans le tube Eppendorf pendant environ 1 sec. L’aiguille se remet ensuite en place.
7. Lorsque le plateau est atteint, la valeur “+0000” doit s’afficher. Si la valeur “-0000” s’affiche, l’indicateur d’erreur réagit et la sonnerie retentit qui peut être arrêtée en appuyant sur le bouton vert.
8. Relever la tête de mesure.
ATTENTION: lorsque l’échantillon est totalement congelé, il faut d’abord réchauffer le tube Eppendorf avec les doigts avant de le retirer.
9. Enlever le godet.
Nettoyer la thermistance à l’aide d’un papier ménage humide sans la casser et en évitant absolument de faire pénétrer de l’eau dans le puit du bloc métallique réfrigéré, ce qui entraînerait la formation de glace. Dans ce cas, débrancher l’appareil pendant 2 min environ, puis sécher l’ouverture du bloc à l’aide d’un tampon de coton.
10 Pipeter 100 μl de la solution standard à 500 mOsm / kg dans un tube Eppendorf.
11 Procédez selon les points 3 à 7.
12 Lorsque le plateau est atteint, la valeur de “500” s’affiche à l’écran.
13 Procédez ensuite selon les points 8 et 9.
14 Pour la mesure de vos échantillons, procédez comme précédemment depuis le point 10 (il n'est bien sûr plus nécessaire de mesurer le point de congélation de l'eau).
Les lambeaux épidermiques, constitués d'une assise cellulaire unique, permettent une observation microscopique aisée des cellules qui les composent. Les vacuoles de ces tissus occupent la presque totalité de l'espace cellulaire. Il est ainsi possible, en plongeant ces lambeaux dans des solutions de potentiel osmotique variable, d'étudier les échanges d'eau entre les cellules et le milieu extérieur, et, d'obtenir, sur la base d'observations microscopiques, des informations sur la potentiel hydrique cellulaire.
- Inciser délicatement, à l'aide d'une lame de rasoir, un carré de 3 mm de côté environ, dans l'épiderme intérieur d'une écaille d'un bulbe d'oignon.
- Détacher avec une paire de brucelles à pointes fines, le fragment épidermique ainsi délimité en le saisissant par un des coins.
- Placer le lambeau (face déchirée vers le bas) entre lame et lamelle dans une goutte de solution d'observation.
- Choisir au microscope une zone facilement observable et examiner après 3-5 min les vacuoles. Etablir, pour chaque solution d'observation utilisée, l'état de plasmolyse des cellules.
A l'isotonie, le potentiel hydrique de la cellule est égal au potentiel osmotique du milieu extérieur.
- On utilisera des solutions de sorbitol (substance qui ne pénètre pas ou peu dans les cellules) à différentes concentrations (1; 5; 10 et 15 %).
- mesurer l'osmolalité de ces solutions avec l'osmomètre à point de congélation
- En travaillant chaque fois avec un lambeau frais, noter la concentration C, pour laquelle il y a une légère plasmolyse. On appellera C ' la concentration qui lui est immédiatement inférieure.
- Préparer alors entre les concentrations C et C ', trois dilutions de sorbitol, en calculant chaque fois les concentrations en %.
- mesurer l'osmolalité de ces solutions avec l'osmomètre à point de congélation
- Refaire pour ces dilutions les mêmes observations que précédemment.
- Déterminer la concentration molaire de sorbitol qui se rapproche le plus de l'isotonie et déduire de l'osmolalité de ces solutions, le potentiel hydrique cellulaire.
- Soit CS la concentration molaire de sorbitol assurant l'isotonie. En supposant que le KNO3 en solution se dissocie complètement, calculer la concentration CK de ce sel, permettant d'obtenir une osmolalité égale à celle du sorbitol à une concentration CS.
- Faire une série d'essais comparables à ceux réalisés précédemment mais avec du KNO3, dont les concentrations limites pourraient être 0,5.CK et 10.CK.
- mesurer l'osmolalité de ces solutions avec l'osmomètre à point de congélation
- Déterminer la concentration CK' de KNO3 entraînant l'isotonie.
- Comparer les deux concentrations CK et CK' et expliquer les différences obtenues.
- Déterminer la valeur du degré d'ionisation du KNO3.
Les tubercules de pommes de terre sont constitués essentiellement de parenchyme de réserve. En mettant à profit la grande homogénéité de ce tissu, il est possible de réaliser un certain nombre d'expériences, telles que la mesure du poids frais ou d'une longueur, sur des fragments de tubercule.
- Laver trois pommes de terre, les éplucher et les placer dans un récipient plein d'eau. Découper les tubercules en parallélépipèdes de section carrée (0.5 cm de côté).
- Après les avoir lavés, conserver les segments de tissus dans un papier filtre humide.
- Ajuster alors les fragments de tissus à une longueur déterminée (Lo), en supprimant les bords inférieurs et supérieurs.
- Essuyer légèrement les segments de tissus ainsi obtenus sur un papier filtre et les répartir en lots de 5.
- Déterminer rapidement le poids frais (PFo) de chaque lot (auquel on attribuera un numéro) et les immerger dans les solutions suivantes :
- Préparer dans 7 béchers des solutions de saccharose, à partir d'une solution stock (2M), selon le tableau suivant (valeurs en ml):
|
bécher |
Solution |
H2O |
Concentration |
|
1 |
0 |
20 |
0.00 |
|
2 |
0,5 |
19,5 |
0,05 |
|
3 |
1 |
19 |
0,10 |
|
4 |
2 |
18 |
0,20 |
|
5 |
4 |
16 |
0,40 |
|
6 |
8 |
12 |
0,80 |
|
7 |
20 |
0 |
2,00 |
- mesurer l'osmolalité de ces solutions avec l'osmomètre à point de congélation
- Après 1 heure, mesurer pour chacun des lots :
- le poids frais PFf, après avoir légèrement essuyé les segments sur un papier filtre.
- la longueur moyenne Lf de chaque lot.
- On calculera alors :
ΔL (%) = ((Lf - Lo) / Lo) . 100
ΔPF (%) = ((PFf - PFo) / PFo) . 100
Reporter sur un graphique, les courbes correspondant au ΔL et au ΔPF, en fonction de la concentration molaire (M) du saccharose.
Présenter un graphique identique, mais cette fois-ci avec l'osmolalité en abscisse.
Pour chacune des courbes, déterminer la valeur de la concentration de saccharose correspondant à l'intersection avec l'abscisse. Calculer à partir de cette valeur celle du potentiel hydrique des tissus.
Exprimer alors celui-ci en MPa.
Le protoplaste est une cellule dont on a fait disparaître (mécaniquement ou enzymatiquement) la paroi. En l'absence des potentiels hydrostatiques (pressions) normalement exercées par la paroi sur le contenu cellulaire, il n'est stable que dans des conditions isotoniques (ou légèrement hypertoniques) et prend alors une forme sphérique. Les protoplastes permettent d'étudier un certain nombre de caractéristiques du plasmalemme (perméabilité, charges, ...). Nous nous bornerons ici à observer la pénétration de deux types de colorants, pour lesquels le plasmalemme présente une perméabilité sélective.
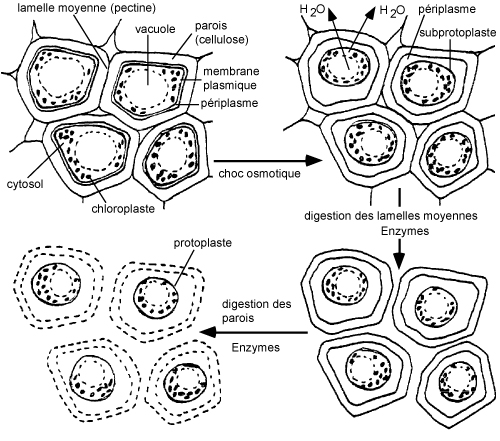
- Pipeter dans une boîte de Pétri 10 ml d'une solution enzymatique déjà prête, contenant des cellulases, des pectinases et du sorbitol. Expliquer le rôle de ces divers composés.
- mesurer l'osmolalité de cette solution avec l'osmomètre à point de congélation
- Peler délicatement, à l'aide d'une brucelle à pointes fines, l'épiderme inférieur de 2-3 feuilles de rampon (Valerianella olitoria) ou de dents-de-lion (Taraxacum sp.), et découper le mésophylle en carrés de 1 cm2 environ. Travailler sur un papier filtre humide, pour éviter le dessèchement.
- Déposer les fragments de feuilles (face pelée vers le bas) à la surface de la solution d'incubation.
- Envelopper la boîte de Pétri d'une feuille d'aluminium et incuber à 20-30 °C durant 30 minutes sur un agitateur orbital (50- 100 tpm).
- Filtrer la suspension de protoplastes à travers un filtre nylon (50-100 μm), pour éliminer les restes de feuilles non digérées.
- Centrifuger la suspension de protoplastes (tubes coniques, 500 tpm, 5 min).
- Eliminer le surnageant. Le culot de protoplastes sera suspendu dans 2-5 ml de solution TL (sorbitol 10%; tampon MES- NaMES ; pH 6.15; 0,05 M).
Mesurer l'osmolalité de cette solution avec l'osmomètre à point de congélation
- Conserver cette suspension de protoplastes dans un erlenmeyer de 50 ml sous agitation modérée.
- Prélever une goutte de suspension de protoplastes et la déposer dans une cellule de comptage. Estimer le nombre de protoplastes isolés et leur concentration (protoplastes / ml).
- Observer alors, à un grossissement de 400 x, les détails morphologiques des protoplastes. Déterminer la position des chloroplastes, des vacuoles, du noyau, et - en fermant le diaphragme pour augmenter la résolution - essayer d'observer la cyclose.
- Ajouter alors sur les bords de la lame de comptage, 1-2 gouttes d'eau. Observer le gonflement des protoplastes et leur éclatement.
- Pipeter délicatement dans 2 tubes à centrifuger (1 et 2), 1 ml de la suspension de protoplastes.
- Ajouter dans les tubes 1 et 2 respectivement 1 goutte des colorants suivants :
I : Rouge neutre 0.1 %
II : Phénosafranine 0.1 %
- Après 5 min, centrifuger et suspendre les protoplastes dans 1 ml de solution TL.
- Pour les deux essais, déterminer, à l'aide de la lame de comptage, le nombre de protoplastes colorés.
- Commenter les résultats.
Il est possible de suivre la pénétration de certains composés à l'intérieur des cellules, en utilisant des substances radioactives. Nous allons ici considérer le cas particulier du ion chlorure.
Les substances radioactives présentent un certain nombre de dangers que les mesures suivantes permettent d'éviter, si elle sont suivies rigoureusement.
Le chlore-36 utilisé lors de ces essais est un émetteur β- d'énergie relativement faible. Ce rayonnement est complètement absorbé par les récipients en plastique ou en verre, Il n'y a donc aucun danger d'irradiation.
1. Recouvrir la place de travail de feuilles de papier de protection spécial et délimiter clairement - à l'aide d'un papier collant de couleur - le champ dans lequel le matériel radioactif sera manipulé. On choisira un endroit calme, en interdisant, si besoin est, les allées et venues.
2. Marquer clairement (stylo feutre, étiquettes ...) toute la verrerie et ustensiles qui seront manipulés dans le champ délimité. Aucun ustensile non marqué ne doit pouvoir y être utilisé.
3. Mettre une blouse et des gants.
4. Faire une fois toutes les manipulations avec une solution non- radioactive.
5. Disposer alors tous les ustensiles de manière à pouvoir les utiliser de manière rationnelle.
6. Ne débuter l'expérience que lorsque chacune des étapes aura été parfaitement assimilée.
7. Tout transfert de solution radioactive doit se faire avec une seringue à tuberculine. Il ne faut jamais pipeter ces solutions avec la bouche.
8. Une fois l'expérience terminée :
- récupérer toutes les solutions dans le récipient mis à disposition.
- déposer les ustensiles (brucelles, ...) dans le bac de décontamination.
- déposer les papier filtres, aluminium, boîtes de Pétri, pointes de pipettes, ... dans un sac en aluminium.
- laver la place de travail avec une solution de NaCl et la rincer soigneusement.
- déposer les gants dans le sac en aluminium.
9. En cas d'incident, avertir immédiatement l'assistant.
L'expérience sera faite sur des disques de tubercules de pomme de terre.
Préparer 60 disques (aussi homogènes que possible) et les mettre en pré-incubation dans 10 ml de tampon (Mes-NaMes; 0,05M; pH 6.15; 30 min; agitateur orbital 100 tpm).
Schéma de l'expérience
- Remplir 14 tubes en plastique de scintillateur avec la pipette automatique Socorex, les boucher et les déposer sur le support ad hoc.
- Arrêter la pré-incubation et récupérer les disques dans un tamis. Après les avoir rapidement séchés en surface, les déposer dans un boîte de Pétri en plastique. Disposer cette dernière dans une grande boîte de Pétri (30 cm) en verre.
- Au temps t=0 (chronomètre), mettre les disques en présence de 10 ml de solution radioactive (tampon + 36Cl-). A l'aide d'une brucelle, prélever immédiatement
(t < 10 sec) 3 disques, que l'on rincera abondamment à l'eau distillée et que l'on déposera dans un petit tube à scintillation. Boucher le tube et l'agiter au Vortex pendant 10 sec, puis le mettre sur le support.
- Enclencher l'agitation (50-100 tpm).
- Toutes les 6 min (ou selon les indications de l'assistant), durant 36 min, arrêter l'agitation et faire un prélèvement du matériel.
- A la fin de l'incubation, récupérer les disques restants sur un tamis et les rincer avec 20 ml de tampon.
- Eliminer la solution dans un flacon mis à disposition.
- Incuber, dans une nouvelle boîte de Pétri, les disques restants en présence de 10 ml de tampon.
- Faire de nouveaux prélèvements de matériel (comme ci-dessus) durant 30 min.
- Bien agiter tous les tubes au vortex et les placer dans des flacons à scintillation, en suivant les indications de l'assistant.
Déposer les flacons numérotés dans un compteur à scintillation. Après 30 min d'équilibration de la température, on pourra faire un premier comptage. Selon la valeur du rendement, agiter tous les flacons et faire une deuxième comptage.
Cette opération sera effectuée par l'assistant, qui donnera ultérieurement les résultats des comptages.
Reporter sur un graphique les cpm (coups/min), en fonction du temps.
Discuter les résultats.