| |
|
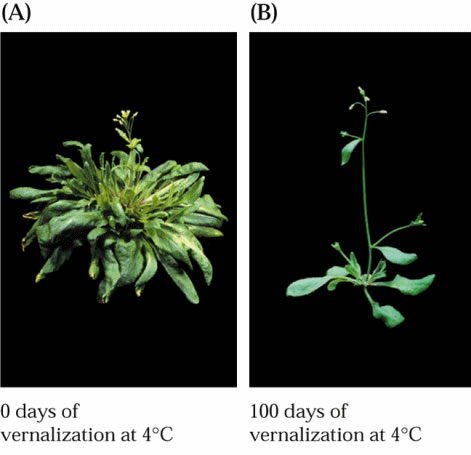
 La
spirale phyllotactique est très
visible chez les composées (ici le tournesol). Le film
(format *.avi 2.5 Mb) suivant vous montre l'effet d'une modification
de l'angle qui sépare deux primordia (normalement 137.5 °) sur
la structure d'une plante de cotton. Dans ce cas l'angle de optimal a aussi
comme effet de maximiser l'interception du rayonnement solaire. La théorie
mathématique de la phyllotaxie est assez complexe vous pouvez essayer
de vous familiariser en consultant le site suivant: http://www.math.smith.edu/~phyllo
La
spirale phyllotactique est très
visible chez les composées (ici le tournesol). Le film
(format *.avi 2.5 Mb) suivant vous montre l'effet d'une modification
de l'angle qui sépare deux primordia (normalement 137.5 °) sur
la structure d'une plante de cotton. Dans ce cas l'angle de optimal a aussi
comme effet de maximiser l'interception du rayonnement solaire. La théorie
mathématique de la phyllotaxie est assez complexe vous pouvez essayer
de vous familiariser en consultant le site suivant: http://www.math.smith.edu/~phyllo
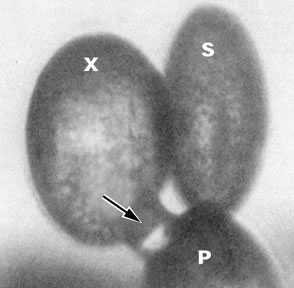
 La
germination du grain de pollen s'accompagne
d'une redistribution des constituants cellulaire. Un dépôt
de callose
confine la partie apicale métaboliquement active du tube pollinique.
Ce dépôt fluorescent est visible sur la photographie prise
en lumière UV.
La
germination du grain de pollen s'accompagne
d'une redistribution des constituants cellulaire. Un dépôt
de callose
confine la partie apicale métaboliquement active du tube pollinique.
Ce dépôt fluorescent est visible sur la photographie prise
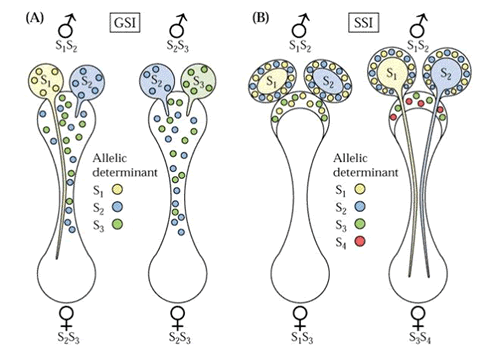
en lumière UV. Au
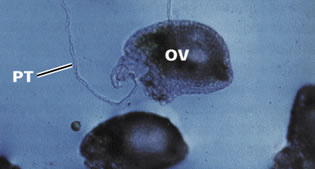
contact du stigmate le grain de pollen peut germer sauf s'il y a incompatibilité
sporophytique. La parois
d'exine du grain de pollen est d'origine sporophytique; elle est responsable
de l'extrème variété morphologique des grains de pollen
et de l'incompatibilité de type sporophytique.
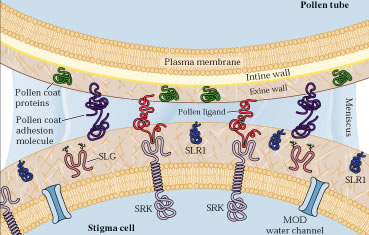
Les protéines impliquées dans la reconnaissance sont un ligand
présent dans l'exine et un recepteur (SRK S locus receptor kinase)
situé à la surface des cellules du stigmate. L'adhésion
des grains de pollen est facilitée par la présence de proteines
d'adhésion et de glycoprotéines (SLG)
Au
contact du stigmate le grain de pollen peut germer sauf s'il y a incompatibilité
sporophytique. La parois
d'exine du grain de pollen est d'origine sporophytique; elle est responsable
de l'extrème variété morphologique des grains de pollen
et de l'incompatibilité de type sporophytique.
Les protéines impliquées dans la reconnaissance sont un ligand
présent dans l'exine et un recepteur (SRK S locus receptor kinase)
situé à la surface des cellules du stigmate. L'adhésion
des grains de pollen est facilitée par la présence de proteines
d'adhésion et de glycoprotéines (SLG) Développement
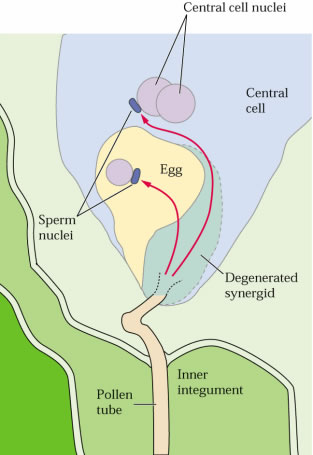
de l'embryon d'Arabidopsis thaliana.
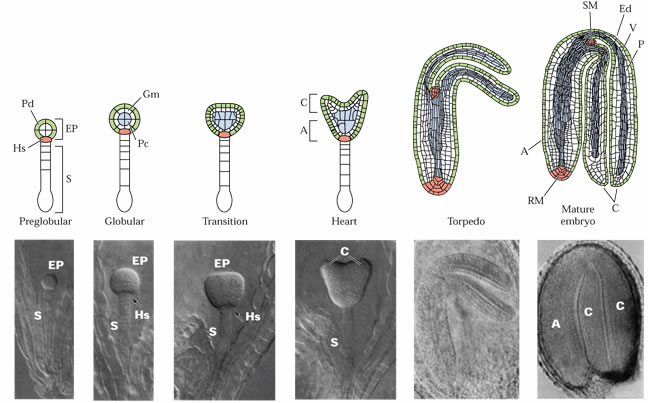
Développement
de l'embryon d'Arabidopsis thaliana.
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}