Hormones
végétales 
B.
Le rôle des hormones dans le développement des végétaux
Auxines
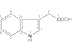 Les
auxines
ont été parmi les premières phytohormones
caractérisées (AIA : acide b-indolyl-acétique - hétéroauxine).
Les
auxines
ont été parmi les premières phytohormones
caractérisées (AIA : acide b-indolyl-acétique - hétéroauxine).
- Les auxines de synthèses
ont été produites dès la moitié du 20ème
siècle; ce sont en particulier l'acide 2,4-dichloro-phénoxyacétique
(2,4-D) et acide alpha-naphtyl-acétique)
- Les auxines sont impliquées
dans le phototropisme;
c'est d'ailleurs à la suite des expériences sur le phototropisme
de Charles Darwin, publiées dans son livre "The
Power of Movement in Plants" (ou document *.rtf
384 kb), que l' implication d' une transmission de l' information à
distance dans les tissus a été formulée. Dans ce cas
il est démontré que les auxines stimulent l' élongation
(auxésis) des organes aériens (tiges)
- L'auxine agit sur la croissance
des racines en l' inhibant
à faible concentration déjà , c' est ce phénomène
qui est mis en jeu dans le géotropisme.
- L'auxine est produite dans les
bourgeons et transportés de manière basipète (vers
la base de la plante); elle responsable de la dominance apicale, c'est à
dire de l'inhibition de croissance des bourgeons latéraux.
- Le transport polarisé
de l'auxine est du à la distribution asymétrique de transporteurs
spécifiques; ce transport peut être inhibé par une anti-auxine,
le 2,3,5-tri-iodobenzoate (TIBA).
- L'auxine stimule la rhizogenèse
(l'apparition de racines sur les tiges et autres organes) et se trouve donc
être l'hormone de bouturage par excellence.
- L'auxine est impliquée
dans le développement des fruits (parthénocarpie) et dans
le phénomène d'abscission qui précède la chute
des feuilles.
- L'auxine peut agir sur la détermination
du sexe chez les cucurbitacées monoïques
en favorisant la production de fleurs femelles (peut-être par l'intermédiaire
de l'éthylène).
- L'auxine est aussi utilisée
pour stimuler l'embryogenèse à partir des
microspores (production de plantes haploïdes)
- L'auxine agit à l'extérieur
de la cellule en se fixant au niveau de la membrane plasmique (récepteur
ABP1 "Auxin-binding protein 1"), son action s'accompagne d'une
acidification de la paroi cellulaire qui contribue à l'extension
de celle ci.
- Une auxine comme l'AIA peut
aussi agir au niveau de l'expression génétique en se liant
à des protéines cytosolique ARF ("auxin response factor).
ARF1 est un facteur de transcription, il peut entrer dans le noyau et se
lier à l'ADN sur une séquence
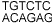 .
Cette séquence ("auxine response element") se retrouve
dans les séquences promoteur des gènes dont la transcription
est affectée par l'auxine.
.
Cette séquence ("auxine response element") se retrouve
dans les séquences promoteur des gènes dont la transcription
est affectée par l'auxine.
- Les auxines de synthèse
peuvent fonctionner comme herbicide (2,4-D). Elles ont été
utilisées comme agent défoliant (en particulier l'auxine 2,4,5-T
dont les dérivés sont tératogènes) par l'armée
des Etats-Unis lors de la guerre du Vietnam. Cette utilisation a fait l'objet
d'une condamnation par le "International
War Crimes Tribunal on Vietnam" en 1967
Cytokinines
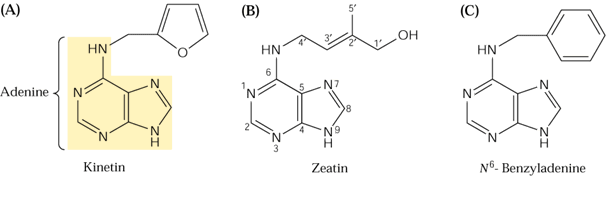
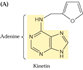 La
première cytokinine identifiée, la kinétine,
est un produit de décomposition de l'ADN par la chaleur. La zéatine
est une cytokinine naturelle et la BAP (benzyl-adénine) une cytokinine
de synthèse
La
première cytokinine identifiée, la kinétine,
est un produit de décomposition de l'ADN par la chaleur. La zéatine
est une cytokinine naturelle et la BAP (benzyl-adénine) une cytokinine
de synthèse- Les cytokinines
sont synthétisées
à partir de l'AMP et du diméthyl-allyl-pyrophosphate (le précurseur
des isoprénoïdes, cf. chapitre de ce cours sur le métabolisme
secondaire). Un équilibre
précis entre leur accumulation et leur dégradation est essentiel
au bon développement des plantes.
- Les cytokinines sont impliquées
avec les auxines dans le développement de la technique de culture
de tissus et dans le procédé industriel de la micropropagation
végétative. Un équilibre
correct entre la concentration d'auxine (ici l'IBA ou acide indolyl-butyrique)
et celle de cytokinine (ici la zéatine) est essentiel.
Gibbérellines
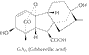 Les
gibberellines
ont été découvertes en 1926 au Japon lors d'études
sur le champignon pathogène Gibberella fujikuroï qui
cause une maladie du riz (bakanae) caractérisée par une élongation
anormale des entre-noeuds. Leur structure
fut élucidée en 1956 seulement; il existe actuellement plus
de 80 gibbérellines différentes.
Les
gibberellines
ont été découvertes en 1926 au Japon lors d'études
sur le champignon pathogène Gibberella fujikuroï qui
cause une maladie du riz (bakanae) caractérisée par une élongation
anormale des entre-noeuds. Leur structure
fut élucidée en 1956 seulement; il existe actuellement plus
de 80 gibbérellines différentes. La
synthèse des gibbérellines (voie des diterpènes) implique
l'enzyme clé ent-kaurène-synthase
dont l'absence (mutation) est une des causes du nanisme. Ce nanisme peut
être corrigé par une application
de GA3. Cette enzyme codée par un gène nucléaire
est localisée dans les plastes. Ce gène (Le
,allèle récessif le) est parfois
nommé "gène de Mendel" puisqu'il est responsable
du nanisme du pois observé et étudié par Gregor Mendel
La
synthèse des gibbérellines (voie des diterpènes) implique
l'enzyme clé ent-kaurène-synthase
dont l'absence (mutation) est une des causes du nanisme. Ce nanisme peut
être corrigé par une application
de GA3. Cette enzyme codée par un gène nucléaire
est localisée dans les plastes. Ce gène (Le
,allèle récessif le) est parfois
nommé "gène de Mendel" puisqu'il est responsable
du nanisme du pois observé et étudié par Gregor Mendel- Les gibbérellines jouent
un rôle clé dans l'induction
de la floraison, ici chez la carotte où elles remplacent un traitement
par le froid (les plantes sans traitement par le froid ou les gibbéréllines
restent à l'état végétatif).
- Les gibbérellines induisent
la synthèse de novo de
l'alpha-amylase dans la couche à aleurone (-2)
des caryopses de céréales (la beta-amlylase est, elle, simplement
activée à partir d'une protéine pro-enzymatique pré-existante
(-3 ).
- Les antigibbérellines
inactivent la ent-kaurène-synthase et empêchent ainsi
la synthèse des gibbéréllines dans les plantes traitées
(induction du nanisme)
Ethylène
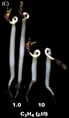 L'éthylène
cause des malformations bien visibles des jeunes
germinations de pois pour la première fois observées par
Neliubov en 1886 (à St-Peterbourg) dans une étude sur les
effets du gaz de ville. On constate bien le dia-géotropisme, l'inhibition
de la croissance et le gonflement des tissus (réponse
triple) causé par une augmentation de la teneur en éthylène.
L'éthylène
cause des malformations bien visibles des jeunes
germinations de pois pour la première fois observées par
Neliubov en 1886 (à St-Peterbourg) dans une étude sur les
effets du gaz de ville. On constate bien le dia-géotropisme, l'inhibition
de la croissance et le gonflement des tissus (réponse
triple) causé par une augmentation de la teneur en éthylène.- L'éthylène
est synthétisé à partir de la S-adénosine-méthionine
et par l'intermédiaire de l'ACC (Acide 1-amino-propane-carboxylique)
- L'éthylène joue
un rôle capital dans le mûrissement
des fruits en stimulant la synthèse des enzymes lytiques (par
exemple les pectinases). Les plantes qui sont devenues
insensibles à l'éthylène suite à une mutation
(par transgenèse TR) présentent des phénotypes
caractéristiques: élongation excessive (A),
retard ou absence de maturation des fruits (B & C),
fleurs de longue durée ne flétrissant plus (D)
et qui présentent un intérêt incontestable pour la fleur
coupée.
L'acide abscissique
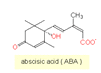 L'acide
abscissique (ABA) est (comme les gibbérellines) produit dans
la voie de biosynthèse des terpènes (sesquiterpènes).
L'acide
abscissique (ABA) est (comme les gibbérellines) produit dans
la voie de biosynthèse des terpènes (sesquiterpènes).- L'ABA est impliqué dans
les mécanismes d'ouverture
et de fermeture des stomates. Le dérivé de l'ABA : 1-(2-nitrophényl)éthyl-ABA
est transformé en ABA sous l'effet de la lumiére UV. On peut
charger ce dérivé dans des cellules de garde puis activer
la formation de l'ABA par une irradiation UV; dans ce cas les stomates
s'ouvrent prouvant ainsi le rôle direct de l'ABA dans les mécanismes
d'ouverture.
- Les mutants de maïs insensibles
à l'ABA montrent un phénotype
vivipare (vp1). Ce phénotype est dû au fait que l'entrée
en dormance de l'embryon ne se fait plus en l'absence de l'hormone de dormance
(ABA) et que cet embryon germe immédiatement sur la plante mère.
- L'importance de l'abscission
dans la domestication des espèces végétales (en
particulier des céréales) est cruciale
Les brassinostéroïdes
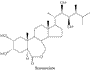 Les
brassinostéroïdes sont des stérols
(voie des triterpènes) qui furent caractérisés, en
particulier, par des chercheurs japonais utilisant un test
biologique sur la feuille de riz. Ce test mesure l'angle d'inclinaison
de la feuille par rapport à la gaine foliaire (cette inclinaison
croissante sous l'effet des brassinostéroïdes est due au gonflement
anormal des cellules adaxiales).
Les
brassinostéroïdes sont des stérols
(voie des triterpènes) qui furent caractérisés, en
particulier, par des chercheurs japonais utilisant un test
biologique sur la feuille de riz. Ce test mesure l'angle d'inclinaison
de la feuille par rapport à la gaine foliaire (cette inclinaison
croissante sous l'effet des brassinostéroïdes est due au gonflement
anormal des cellules adaxiales).- Les mutants
d'Arabidopsis déficients dans la synthèse des brassinostéroïdes
ont un phénotype très clair avec un déficit manifeste
de la croissance.

 La
première cytokinine identifiée, la kinétine,
est un produit de décomposition de l'ADN par la chaleur. La zéatine
est une cytokinine naturelle et la BAP (benzyl-adénine) une cytokinine
de synthèse
La
première cytokinine identifiée, la kinétine,
est un produit de décomposition de l'ADN par la chaleur. La zéatine
est une cytokinine naturelle et la BAP (benzyl-adénine) une cytokinine
de synthèse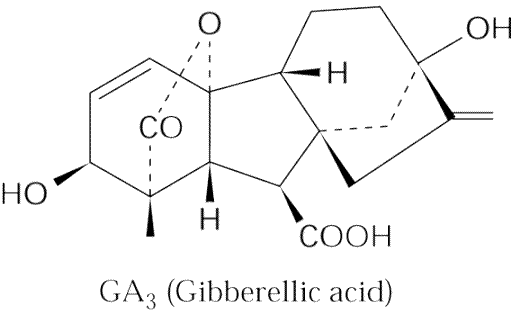 Les
gibberellines
ont été découvertes en 1926 au Japon lors d'études
sur le champignon pathogène Gibberella fujikuroï qui
cause une maladie du riz (bakanae) caractérisée par une élongation
anormale des entre-noeuds. Leur structure
fut élucidée en 1956 seulement; il existe actuellement plus
de 80 gibbérellines différentes.
Les
gibberellines
ont été découvertes en 1926 au Japon lors d'études
sur le champignon pathogène Gibberella fujikuroï qui
cause une maladie du riz (bakanae) caractérisée par une élongation
anormale des entre-noeuds. Leur structure
fut élucidée en 1956 seulement; il existe actuellement plus
de 80 gibbérellines différentes. La
synthèse des gibbérellines (voie des diterpènes) implique
l'enzyme clé ent-kaurène-synthase
dont l'absence (mutation) est une des causes du nanisme. Ce nanisme peut
être corrigé par une application
de GA3. Cette enzyme codée par un gène nucléaire
est localisée dans les plastes. Ce gène (Le
,allèle récessif le) est parfois
nommé "gène de Mendel" puisqu'il est responsable
du nanisme du pois observé et étudié par Gregor Mendel
La
synthèse des gibbérellines (voie des diterpènes) implique
l'enzyme clé ent-kaurène-synthase
dont l'absence (mutation) est une des causes du nanisme. Ce nanisme peut
être corrigé par une application
de GA3. Cette enzyme codée par un gène nucléaire
est localisée dans les plastes. Ce gène (Le
,allèle récessif le) est parfois
nommé "gène de Mendel" puisqu'il est responsable
du nanisme du pois observé et étudié par Gregor Mendel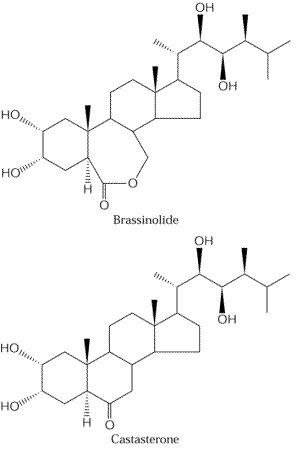 Les
brassinostéroïdes sont des stérols
(voie des triterpènes) qui furent caractérisés, en
particulier, par des chercheurs japonais utilisant un test
biologique sur la feuille de riz. Ce test mesure l'angle d'inclinaison
de la feuille par rapport à la gaine foliaire (cette inclinaison
croissante sous l'effet des brassinostéroïdes est due au gonflement
anormal des cellules adaxiales).
Les
brassinostéroïdes sont des stérols
(voie des triterpènes) qui furent caractérisés, en
particulier, par des chercheurs japonais utilisant un test
biologique sur la feuille de riz. Ce test mesure l'angle d'inclinaison
de la feuille par rapport à la gaine foliaire (cette inclinaison
croissante sous l'effet des brassinostéroïdes est due au gonflement
anormal des cellules adaxiales).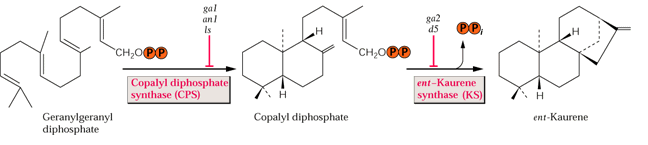{kind=link}
{kind=link}
{kind=link}