Biologie
du stress 
G. La biologie
du "stress"
Les
stress biotiques et abiotiques combinés sont responsable de pertes
considérable dans les cultures; ces pertes peuvent atteindre le 80% du
rendement optimal.
Stress hydriques
- Le manque d'eau (irrigation déficiente
ou nappe phréatique inaccessible) et la rareté des précipitations
sont les causes principale du stress hydriques (cf chapitre
du cours consacré à l'eau).
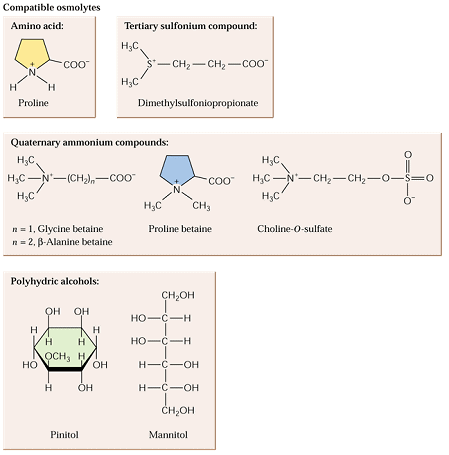
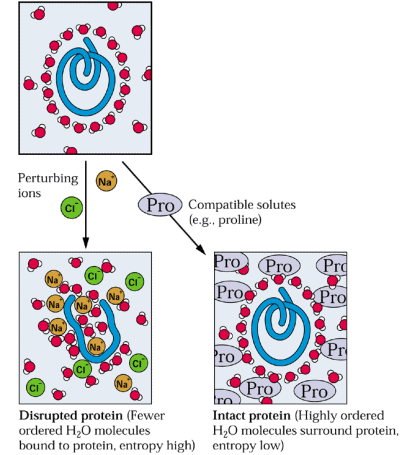
 L'accumulation
de molécules spéciales appelées osmolytes
accompagne le stress hydrique. Ces molécules agissent comme agent de
protection des macromolécules contre
la déshydratation.
L'accumulation
de molécules spéciales appelées osmolytes
accompagne le stress hydrique. Ces molécules agissent comme agent de
protection des macromolécules contre
la déshydratation.
Stress de température
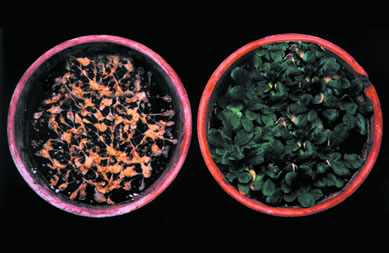
- Les végétaux peuvent
s'adapter à des température extrèmes soit basse ( en
dessous de 12°C) soit élevées (en dessus de 35°C). Ils
sont aussi capable de résister à la congélation (<<
0° C). La résitance au froid peut être acquise par un prétraitement
(adaptation). La plante de droite a été adaptée pendant
4 jours à 4°C avant d'être placée avaec la plante
témoin à -5°C pendant 4 jours.
Stress salin
- La présence de concentrations
de NaCl supérieurs à 50mM dans les sols est, en général,
défavorable à la plupart des espèces végétales
(en particulier celle que l'on regroupe sous le nom de glycophytes. Le NaCl
en lui-même est toxique, mais le stress salin s'accompagne souvent d'une
baisse importante du potentiel hydrique du sol (cf. stress hydrique)
Interactions des plantes avec les
pathogène
-
Le botrytis
cinerea est un champignon nécrotrophe qui attaque les tissus
à l'aide d'enzymes de dégradation de la parois.
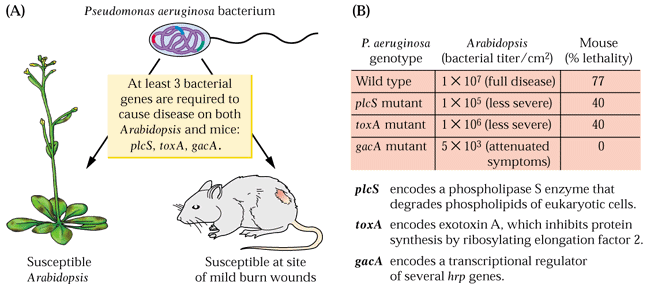
- Les génes nécessaire
à une bactérie pathogéne, à la fois pour l'élicitation
de la réponse hypersensible et pour la pathogénicité
sont appelés gènes "hrp" (Hypersensitive Response
& Pathogenicity). Chez la bactérie Pseudomonas aeruginosa,
les gènes "hrp" codent pour des protéines qui sont
nécessaires à la pathogénicité à la fois
chez Arabidopsis
et chez la souris.
- La réponse hypersensible
est le temoin d'une résistance (incompatibilité) entre le pathogène
et la plante. Ainsi, la bactérie Erwinia amylora cause du
"feux bactérien" qui décime les arbres fruitiers (poirier,
pommier), déclenche une réponse HR chez les plantes non-hôte
comme le tabac par exemple. Chez la tomate, le gène Pto confère
une résistance à Pseudomonas syringae pv tomato avrPto
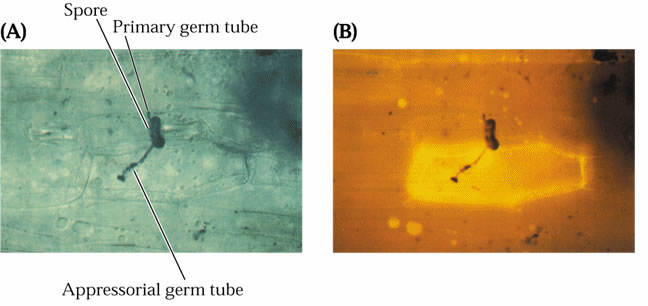
- La reaction hypersensible est
caractérisée par une modification du métabolisme de la
cellule attaquée (dans cette photo elle
devient fluorescente aux UV suite à une attaque par un champignon).
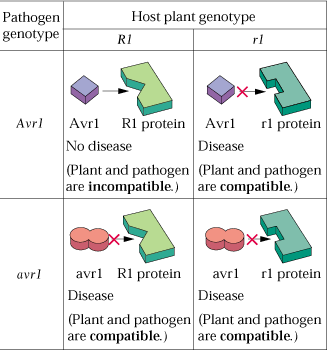
- Le modèle d'interaction
hôte-pathogène appelé "gène
pour gène" implique la présence d'un gène d'avirulence
chez le pathogène et de résistance chez l'hôte. Chez la
tomate par exemple, le gène Pto confère une résistance
à Pseudomonas syringae pv tomato avrPto.
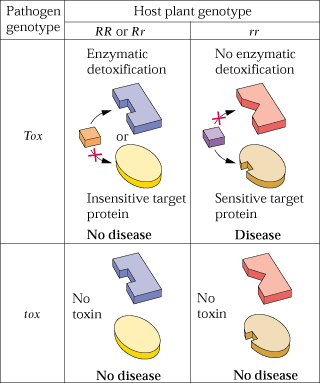
- Certain pathogènes produisent
des toxines spécifiques, dans ce cas le modèle
génétique implique, chez le pathogène, un gène
impliqué dans la synthèse de la toxine et chez l'hôte,
un gène impliqué dans la synthèse d'un récepteur
ou d'une enzyme de détoxification.
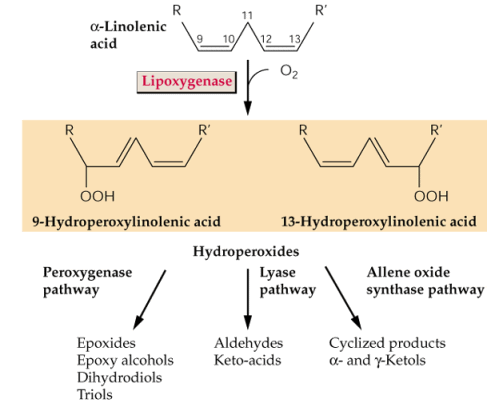
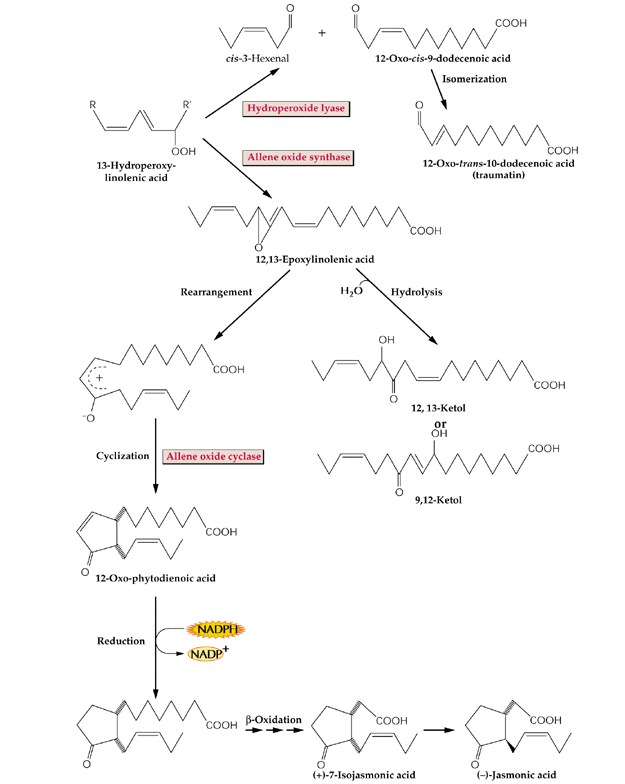
- L'acide linolénique libéré
de la membrane plasmique par l'action d'une phospholipase (suite à
l'action d'un éliciteur par exemple) est modifié par une lipoxygénase
(LOX). C'est une phase essentielle dans la formation des oxylipines
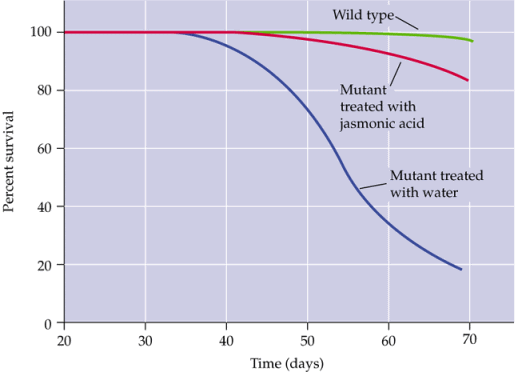
(acide jasmonique). L'acide jasmonique protège les plantes contre les
attaques par des insectes. Des plantes mutantes incapables de synthétiser
du jasmonate ne résistent pas, à moins de subir un traitement
par de l'acide jasmonique.
Phytoalexines
Virus

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}